![]() –†–Ķ–Ņ–Ľ–ł–ļ–į—Ü–ł—ė–į –Ĺ–į –Ē–Ě–ö. –Ē–≤–ĺ—ė–Ĺ–ĺ—Ā–Ņ–ł—Ä–į–Ľ–Ĺ–į—ā–į –ľ–ĺ–Ľ–Ķ–ļ—É–Ľ–į –Ĺ–į –Ē–Ě–ö —Ā–Ķ –ĺ–ī–ľ–ĺ—ā—É–≤–į –ł —Ā–Ķ–ļ–ĺ—ė–į –ĺ–ī–ľ–ĺ—ā–į–Ĺ–į –≤–Ķ—Ä–ł–≥–į —Ā–Ľ—É–∂–ł –ļ–į–ļ–ĺ —ą–į–Ī–Ľ–ĺ–Ĺ –∑–į —Ā–ł–Ĺ—ā–Ķ–∑–į –Ĺ–į –Ĺ–ĺ–≤–į, –ĺ—Ā–Ĺ–ĺ–≤–į—ė—ú–ł —Ā–Ķ –Ĺ–į –ļ–ĺ–ľ–Ņ–Ľ–Ķ–ľ–Ķ–Ĺ—ā–į—Ä–Ĺ–ĺ—Ā—ā–į –Ĺ–į –Ī–į–∑–Ĺ–ł—ā–Ķ –Ņ–į—Ä–ĺ–≤–ł.
–†–Ķ–Ņ–Ľ–ł–ļ–į—Ü–ł—ė–į –Ĺ–į –Ē–Ě–ö. –Ē–≤–ĺ—ė–Ĺ–ĺ—Ā–Ņ–ł—Ä–į–Ľ–Ĺ–į—ā–į –ľ–ĺ–Ľ–Ķ–ļ—É–Ľ–į –Ĺ–į –Ē–Ě–ö —Ā–Ķ –ĺ–ī–ľ–ĺ—ā—É–≤–į –ł —Ā–Ķ–ļ–ĺ—ė–į –ĺ–ī–ľ–ĺ—ā–į–Ĺ–į –≤–Ķ—Ä–ł–≥–į —Ā–Ľ—É–∂–ł –ļ–į–ļ–ĺ —ą–į–Ī–Ľ–ĺ–Ĺ –∑–į —Ā–ł–Ĺ—ā–Ķ–∑–į –Ĺ–į –Ĺ–ĺ–≤–į, –ĺ—Ā–Ĺ–ĺ–≤–į—ė—ú–ł —Ā–Ķ –Ĺ–į –ļ–ĺ–ľ–Ņ–Ľ–Ķ–ľ–Ķ–Ĺ—ā–į—Ä–Ĺ–ĺ—Ā—ā–į –Ĺ–į –Ī–į–∑–Ĺ–ł—ā–Ķ –Ņ–į—Ä–ĺ–≤–ł.
–†–Ķ–Ņ–Ľ–ł–ļ–į—Ü–ł—ė–į –ł–Ľ–ł —É–ī–≤–ĺ—ė—É–≤–į—ö–Ķ (–ĺ–ī replica —ą—ā–ĺ –∑–Ĺ–į—á–ł –Ņ—Ä–ł–ľ–Ķ—Ä–ĺ–ļ) ‚ÄĒ –Ņ—Ä–ĺ—Ü–Ķ—Ā –≤–ĺ –≥–Ķ–Ĺ–Ķ—ā–ł–ļ–į—ā–į —Ā–ĺ –ļ–ĺ—ė –ľ–ĺ–Ľ–Ķ–ļ—É–Ľ–į—ā–į –Ĺ–į –Ē–Ě–ö —Ā–Ķ —É–ī–≤–ĺ—ė—É–≤–į. –ě–≤–į–į –į–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā —Ā–Ķ —Ā–Ľ—É—á—É–≤–į –≤–ĺ –ł–Ĺ—ā–Ķ—Ä—Ą–į–∑–į—ā–į –ĺ–ī –ļ–Ľ–Ķ—ā–ĺ—á–Ĺ–ł–ĺ—ā —Ü–ł–ļ–Ľ—É—Ā –ł –Ņ—Ä–Ķ–ļ—É –Ĺ–Ķ–į –ļ–Ľ–Ķ—ā–ļ–ł—ā–Ķ –ļ–ĺ–ł —ú–Ķ —Ā–Ķ –ī–ĺ–Ī–ł—ė–į—ā –ĺ–ī –ļ–Ľ–Ķ—ā–ĺ—á–Ĺ–į—ā–į –ī–Ķ–Ľ–Ī–į –≥–ĺ –Ĺ–į—Ā–Ľ–Ķ–ī—É–≤–į–į—ā —Ü–Ķ–Ľ–ĺ–ļ—É–Ņ–Ĺ–ł–ĺ—ā –≥–Ķ–Ĺ–ĺ–ľ, –ĺ–ī–Ĺ–ĺ—Ā–Ĺ–ĺ —Ü–Ķ–Ľ–ĺ—Ā–Ĺ–ł–ĺ—ā —Ā–Ķ—ā –ĺ–ī –≥–Ķ–Ĺ–ł –Ņ—Ä–ł—Ā—É—ā–Ĺ–ł –≤–ĺ –Ķ–ī–Ķ–Ĺ –ĺ—Ä–≥–į–Ĺ–ł–∑–į–ľ. –ö–į—ė –Ķ—É–ļ–į—Ä–ł–ĺ—ā–ł—ā–Ķ, –ľ–ĺ–Ľ–Ķ–ļ—É–Ľ–ł—ā–Ķ –Ĺ–į –Ē–Ě–ö –ļ–ĺ–ł –≥–ĺ –≥—Ä–į–ī–į—ā –≥–Ķ–Ĺ–ĺ–ľ–ĺ—ā —Ā–Ķ –Ņ–ĺ–≤—Ä–∑–į–Ĺ–ł —Ā–ĺ –Ī–Ķ–Ľ–ļ–ĺ–≤–ł–Ĺ–ł –ł –≤–Ľ–Ķ–≥—É–≤–į–į—ā –≤–ĺ —Ā–ĺ—Ā—ā–į–≤ –Ĺ–į —Ö—Ä–ĺ–ľ–ĺ–∑–ĺ–ľ–ł—ā–Ķ, –ĺ–ī –ļ–ĺ–ł —Ā–Ķ–ļ–ĺ—ė —Ā–ĺ–ī—Ä–∂–ł –Ķ–ī–ł–Ĺ–Ķ—á–Ĺ–į –Ľ–ł–Ĺ–Ķ–į—Ä–Ĺ–į –Ē–Ě–ö-–ľ–ĺ–Ľ–Ķ–ļ—É–Ľ–į. –ē—É–ļ–į—Ä–ł–ĺ—ā—Ā–ļ–ł—ā–Ķ —Ö—Ä–ĺ–ľ–ĺ–∑–ĺ–ľ–ł —Ā–Ķ –Ĺ–į–ĺ—ď–į–į—ā –≤–ĺ –ļ–Ľ–Ķ—ā–ĺ—á–Ĺ–ĺ—ā–ĺ —ė–į–ī—Ä–ĺ. –ď–Ķ–Ĺ–ĺ–ľ–ł—ā–Ķ –Ĺ–į –Ī–į–ļ—ā–Ķ—Ä–ł—Ā–ļ–ł—ā–Ķ –ļ–Ľ–Ķ—ā–ļ–ł (–Ņ—Ä–ĺ–ļ–į—Ä–ł–ĺ—ā–ł), –ļ–į—ė –ļ–ĺ–ł –ĺ—ā—Ā—É—Ā—ā–≤—É–≤–į —ė–į–ī—Ä–ĺ—ā–ĺ, —Ā–Ķ –Ĺ–į—ė—á–Ķ—Ā—ā–ĺ –ļ—Ä—É–∂–Ĺ–ł –Ē–Ě–ö-–ľ–ĺ–Ľ–Ķ–ļ—É–Ľ–ł (–Ĺ–į—Ä–Ķ—á–Ķ–Ĺ–ł –Ņ–Ľ–į–∑–ľ–ł–ī–ł) –ļ–ĺ–ł —Ā–Ķ –Ņ–ĺ–≤—Ä–∑—É–≤–į–į—ā —Ā–ĺ –Ņ–ĺ—Ā–Ķ–Ī–Ĺ–ł —Ā—ā—Ä—É–ļ—ā—É—Ä–ł –≤–ĺ –ļ–Ľ–Ķ—ā–ĺ—á–Ĺ–į—ā–į –ľ–Ķ–ľ–Ī—Ä–į–Ĺ–į (–ĺ–∑–Ĺ–į—á–Ķ–Ĺ–ł –ļ–į–ļ–ĺ –Ķ–Ņ–ł–∑–ĺ–ľ–ł). –ė –Ņ–ĺ–ļ—Ä–į—ė –≥–ĺ–Ľ–Ķ–ľ–į—ā–į –Ķ–≤–ĺ–Ľ—É—Ü–ł—Ā–ļ–į —Ä–į–∑–Ľ–ł–ļ–į –Ņ–ĺ–ľ–Ķ—ď—É –Ņ—Ä–ĺ–ļ–į—Ä–ł–ĺ—ā–ł—ā–Ķ –ł –Ķ—É–ļ–į—Ä–ł–ĺ—ā–ł—ā–Ķ –ļ–ĺ—ė–į —Ā–Ķ –ľ–Ķ—Ä–ł –≤–ĺ –ľ–ł–Ľ–ł–ĺ–Ĺ–ł –≥–ĺ–ī–ł–Ĺ–ł, –ĺ–ī–Ľ–ł–ļ–ł—ā–Ķ –Ĺ–į –Ņ—Ä–ĺ—Ü–Ķ—Ā–ĺ—ā –Ĺ–į —Ä–Ķ–Ņ–Ľ–ł–ļ–į—Ü–ł—ė–į —Ā–Ķ –∑–į—á—É–≤–į–Ĺ–ł –ł —Ā–Ķ –ľ–Ĺ–ĺ–≥—É —Ā–Ľ–ł—á–Ĺ–ł –ļ–į—ė –ī–≤–Ķ—ā–Ķ –≥–ĺ–Ľ–Ķ–ľ–ł –≥—Ä—É–Ņ–ł –Ĺ–į –∂–ł–≤–ł –ĺ—Ä–≥–į–Ĺ–ł–∑–ľ–ł.
–ě–Ņ—ą—ā –ĺ—Ā–≤—Ä—ā
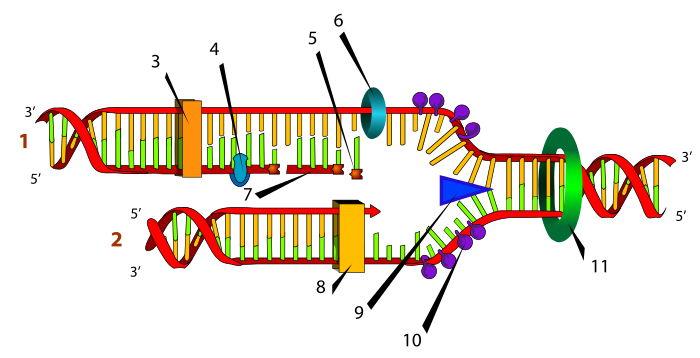
–Ē–Ě–ö-–ľ–ĺ–Ľ–Ķ–ļ—É–Ľ–ł—ā–Ķ –ļ–ĺ–ł –≥–ł –≥—Ä–į–ī–į—ā –≥–Ķ–Ĺ–ĺ–ľ–ł—ā–Ķ –Ĺ–į –Ī–į–ļ—ā–Ķ—Ä–ł–ł—ā–Ķ –ł –Ķ—É–ļ–į—Ä–ł–ĺ—ā—Ā–ļ–ł—ā–Ķ –ļ–Ľ–Ķ—ā–ļ–ł —Ā–Ķ –ī–≤–ĺ–≤–Ķ—Ä–ł–∂–Ĺ–ł –ľ–ĺ–Ľ–Ķ–ļ—É–Ľ–ł –ļ–į—ė –ļ–ĺ–ł —Ā–Ķ–ļ–ĺ—ė–į –≤–Ķ—Ä–ł–≥–į (–ł–Ľ–ł –Ľ–į–Ĺ–Ķ—Ü) —Ā–Ķ —Ā–ĺ—Ā—ā–ĺ–ł –ĺ–ī –Ņ–ĺ–ī–Ķ–ī–ł–Ĺ–ł—Ü–ł –Ĺ–į—Ä–Ķ—á–Ķ–Ĺ–ł –Ĺ—É–ļ–Ľ–Ķ–ĺ—ā–ł–ī–ł. –ě–≤–ł–Ķ –Ĺ—É–ļ–Ľ–Ķ–ĺ—ā–ł–ī–ł –ł–ľ–į–į—ā –Ĺ–į—Ā–ĺ–ļ–į (–Ņ—Ä–į–≤–Ķ—Ü) –Ĺ–į –Ņ—Ä–ĺ—ā–Ķ–≥–į—ö–Ķ, –Ĺ–į —Ā–Ľ–ł—á–Ķ–Ĺ –Ĺ–į—á–ł–Ĺ –ļ–į–ļ–ĺ —ą—ā–ĺ —Ā—ā—Ä–Ķ–Ľ–į—ā–į –ł–ľ–į –≥–Ľ–į–≤–į –ł –ĺ–Ņ–į—ą–ļ–į. –í–ĺ –Ē–Ě–ö-–Ľ–į–Ĺ—Ü–ł—ā–Ķ, –≥–Ľ–į–≤–į—ā–į –Ķ 3‚Ä≤ (—ā—Ä–ł –Ņ—Ä–ł–ľ) –ļ—Ä–į—ė–ĺ—ā –Ĺ–į –Ľ–į–Ĺ–Ķ—Ü–ĺ—ā, –ī–ĺ–ī–Ķ–ļ–į –ĺ–Ņ–į—ą–ļ–į—ā–į –Ķ 5‚Ä≤ (–Ņ–Ķ—ā –Ņ—Ä–ł–ľ). –Ē–≤–Ķ—ā–Ķ –≤–Ķ—Ä–ł–≥–ł –ĺ–ī –Ē–Ě–ö —Ā–Ķ –Ņ—Ä–ĺ—ā–Ķ–≥–į–į—ā –≤–ĺ —Ā–Ņ—Ä–ĺ—ā–ł–≤–Ķ–Ĺ –Ņ—Ä–į–≤–Ķ—Ü –ł —Ā–Ķ –∑–į–ľ–ĺ—ā—É–≤–į–į—ā –Ķ–ī–Ĺ–į –ĺ–ļ–ĺ–Ľ—É –ī—Ä—É–≥–į –≤–ĺ –ī–≤–ĺ—ė–Ĺ–į –∑–į–≤–ĺ—ė–Ĺ–ł—Ü–į (–ī–≤–ĺ—ė–Ĺ–į —Ā–Ņ–ł—Ä–į–Ľ–į), –Ņ—Ä–ł —ą—ā–ĺ —Ā–Ķ –ī—Ä–∂–į—ā –∑–į–Ķ–ī–Ĺ–ĺ —Ā–ĺ –≤–ĺ–ī–ĺ—Ä–ĺ–ī–Ĺ–ł –≤—Ä—Ā–ļ–ł –Ņ–ĺ–ľ–Ķ—ď—É –Ī–į–∑–Ĺ–ł—ā–Ķ –Ņ–į—Ä–ĺ–≤–ł –ĺ–ī –Ĺ—É–ļ–Ľ–Ķ–ĺ—ā–ł–ī–ł—ā–Ķ (–į–ī–Ķ–Ĺ–ł–Ĺ —Ā–Ķ —Ā–Ņ–į—Ä—É–≤–į —Ā–ĺ —ā–ł–ľ–ł–Ĺ, –į –≥–≤–į–Ĺ–ł–Ĺ —Ā–ĺ —Ü–ł—ā–ĺ–∑–ł–Ĺ).
–ó–į –≤—Ä–Ķ–ľ–Ķ –Ĺ–į —Ä–Ķ–Ņ–Ľ–ł–ļ–į—Ü–ł—ė–į—ā–į, –Ľ–į–Ĺ—Ü–ł—ā–Ķ —Ā–Ķ –ĺ–ī–ľ–ĺ—ā—É–≤–į–į—ā —Ā–ĺ –Ņ–ĺ–ľ–ĺ—ą –Ĺ–į –Ķ–Ĺ–∑–ł–ľ –Ĺ–į—Ä–Ķ—á–Ķ–Ĺ –Ē–Ě–ö-—Ö–Ķ–Ľ–ł–ļ–į–∑–į, —ā–į–ļ–į —ą—ā–ĺ –Ĺ–ĺ–≤–į –≤–Ķ—Ä–ł–≥–į –ĺ–ī –Ē–Ě–ö —Ā–Ķ —Ā–ł–Ĺ—ā–Ķ—ā–ł–∑–ł—Ä–į –Ĺ–į —Ā–Ķ–ļ–ĺ—ė–į –ĺ–ī —Ā—ā–į—Ä–ł—ā–Ķ (—ą–į–Ī–Ľ–ĺ–Ĺ—Ā–ļ–ł) –≤–Ķ—Ä–ł–≥–ł –ĺ–ī —Ā—ā—Ä–į–Ĺ–į –Ĺ–į –Ķ–Ĺ–∑–ł–ľ–ĺ—ā –Ē–Ě–ö-–Ņ–ĺ–Ľ–ł–ľ–Ķ—Ä–į–∑–į, –ļ–ĺ—ė –≥–ł –Ņ–ĺ–≤—Ä–∑—É–≤–į –Ĺ–į–ī–ĺ–į—ď–į—á–ļ–ł—ā–Ķ –Ĺ—É–ļ–Ľ–Ķ–ĺ—ā–ł–ī–ł –ĺ–ī —Ü–ł—ā–ĺ–Ņ–Ľ–į–∑–ľ–į—ā–į –≤–ĺ –Ņ–ĺ—Ā–Ľ–Ķ–ī–ĺ–≤–į—ā–Ķ–Ľ–Ĺ–ĺ—Ā—ā (–Ĺ–ł–∑–į) –ļ–ĺ—ė–į, –Ņ–į–ļ, –Ķ –ĺ–ī—Ä–Ķ–ī–Ķ–Ĺ–į –ĺ–ī –Ĺ–ł–∑–į—ā–į –Ĺ–į –Ĺ—É–ļ–Ľ–Ķ–ĺ—ā–ł–ī–ł –Ņ—Ä–ł—Ā—É—ā–Ĺ–ł –Ĺ–į —Ā—ā–į—Ä–į—ā–į, —ą–į–Ī–Ľ–ĺ–Ĺ—Ā–ļ–į –ł–Ľ–ł –ľ–į—ė—á–ł–Ĺ–į –≤–Ķ—Ä–ł–≥–į. –ó–į —Ä–Ķ–Ņ–Ľ–ł–ļ–į—Ü–ł—ė–į—ā–į –Ĺ–į –Ē–Ě–ö —Ā–Ķ –≤–Ķ–Ľ–ł –ī–Ķ–ļ–į –Ķ –Ņ–ĺ–Ľ—É–ļ–ĺ–Ĺ–∑–Ķ—Ä–≤–į—ā–ł–≤–Ĺ–į, –Ī–ł–ī–Ķ—ė—ú–ł —Ā–Ķ–ļ–ĺ—ė–į –ĺ–ī –ī–≤–Ķ—ā–Ķ –ł–ī–Ķ–Ĺ—ā–ł—á–Ĺ–ł —ú–Ķ—Ä–ļ–ł–Ĺ–ł –ľ–ĺ–Ľ–Ķ–ļ—É–Ľ–ł —Ā–ĺ–ī—Ä–∂–ł –Ķ–ī–Ĺ–į –ĺ–ī –ī–≤–Ķ—ā–Ķ –ľ–į—ė—á–ł–Ĺ–ł —ą–į–Ī–Ľ–ĺ–Ĺ—Ā–ļ–ł –≤–Ķ—Ä–ł–≥–ł (—Ā–Ņ—Ä–ĺ—ā–ł–≤–Ĺ–į—ā–į) –ļ–ĺ—ė–į –Ķ —Ā–Ņ–į—Ä–Ķ–Ĺ–į —Ā–ĺ –Ĺ–ĺ–≤–į –≤–Ķ—Ä–ł–≥–į. –ü—Ä–ĺ–ļ–į—Ä–ł–ĺ—ā—Ā–ļ–į—ā–į —Ä–Ķ–Ņ–Ľ–ł–ļ–į—Ü–ł—ė–į –ľ–ĺ–∂–Ķ –ī–į —ā—Ä–į–Ķ —Ā–į–ľ–ĺ –ī–≤–į–Ķ—Ā–Ķ—ā–ł–Ĺ–į –ľ–ł–Ĺ—É—ā–ł, –ī–ĺ–ī–Ķ–ļ–į –Ķ—É–ļ–į—Ä–ł–ĺ—ā—Ā–ļ–į—ā–į –Ķ –∑–Ĺ–į—á–ł—ā–Ķ–Ľ–Ĺ–ĺ –Ņ–ĺ–ī–ĺ–Ľ–≥–į –ł —ā—Ä–į–Ķ –ĺ–ļ–ĺ–Ľ—É –ĺ—Ā—É–ľ —á–į—Ā–į –ļ–į—ė —Ü–ł—Ü–į—á–ł—ā–Ķ.
–ü–ĺ—á–Ķ—ā–Ĺ–į —Ą–į–∑–į –ĺ–ī —Ä–Ķ–Ņ–Ľ–ł–ļ–į—Ü–ł—ė–į—ā–į
–†–Ķ–Ņ–Ľ–ł–ļ–į—Ü–ł—ė–į—ā–į –Ĺ–į –Ē–Ě–ö –∑–į–Ņ–ĺ—á–Ĺ—É–≤–į –Ĺ–į –Ņ–ĺ—Ā–Ķ–Ī–Ĺ–ł –ľ–Ķ—Ā—ā–į –Ĺ–į—Ä–Ķ—á–Ķ–Ĺ–ł –ł–∑–≤–ĺ—Ä–Ĺ–ł –ľ–Ķ—Ā—ā–į (origins) –Ĺ–į –Ē–Ě–ö-—Ä–Ķ–Ņ–Ľ–ł–ļ–į—Ü–ł—ė–į—ā–į. –ē—É–ļ–į—Ä–ł–ĺ—ā—Ā–ļ–ł—ā–Ķ –Ē–Ě–ö-–ľ–ĺ–Ľ–Ķ–ļ—É–Ľ–ł —Ā–ĺ–ī—Ä–∂–į—ā –Ņ–ĺ–≤–Ķ—ú–Ķ —Ä–Ķ–Ņ–Ľ–ł–ļ–į—Ü–ł—Ā–ļ–ł –ł–∑–≤–ĺ—Ä–Ĺ–ł –ľ–Ķ—Ā—ā–į, —Ā–ľ–Ķ—Ā—ā–Ķ–Ĺ–ł –≤–ĺ –ł–Ĺ—ā–Ķ—Ä–≤–į–Ľ–ł –ĺ–ī –ĺ–ļ–ĺ–Ľ—É 100.000 –Ī–į–∑–Ĺ–ł –Ņ–į—Ä–ĺ–≤–ł (100 –ļ–ł–Ľ–ĺ–Ī–į–∑–ł –Ĺ–į –Ņ–į—Ä–ĺ–≤–ł –ł–Ľ–ł 100 kb) –ī–ĺ–Ľ–∂ –Ē–Ě–ö –ľ–ĺ–Ľ–Ķ–ļ—É–Ľ–į—ā–į. –ü–ĺ—Ā—ā–ĺ—ė–į—ā 6 –Ī–ł–Ľ–ł–ĺ–Ĺ–ł –Ī–į–∑–Ĺ–ł –Ņ–į—Ä–ĺ–≤–ł –≤–ĺ —á–ĺ–≤–Ķ—á–ļ–ł–ĺ—ā –≥–Ķ–Ĺ–ĺ–ľ, —Ā–ľ–Ķ—Ā—ā–Ķ–Ĺ–ł –Ĺ–į 46 —Ö—Ä–ĺ–ľ–ĺ–∑–ĺ–ľ–ł, –Ņ–į —Ā–Ņ–ĺ—Ä–Ķ–ī –ĺ–≤–ĺ—ė –Ņ–ĺ–ī–į—ā–ĺ–ļ, —Ā–Ķ–ļ–ĺ—ė —Ö—Ä–ĺ–ľ–ĺ–∑–ĺ–ľ —ú–Ķ –ł–ľ–į –ľ–Ĺ–ĺ–≥—É –ł–∑–≤–ĺ—Ä–Ĺ–ł –ľ–Ķ—Ā—ā–į –∑–į —Ä–Ķ–Ņ–Ľ–ł–ļ–į—Ü–ł—ė–į. –ü—Ä–ĺ–ļ–į—Ä–ł–ĺ—ā—Ā–ļ–ł—ā–Ķ —Ö—Ä–ĺ–ľ–ĺ–∑–ĺ–ľ–ł –Ĺ–į—ė—á–Ķ—Ā—ā–ĺ –ł–ľ–į–į—ā –Ķ–ī–Ĺ–ĺ –ł–∑–≤–ĺ—Ä–Ĺ–ĺ –ľ–Ķ—Ā—ā–ĺ.
–†–Ķ–Ņ–Ľ–ł–ļ–į—Ü–ł—Ā–ļ–ł—ā–Ķ –ł–∑–≤–ĺ—Ä–Ĺ–ł –ľ–Ķ—Ā—ā–į —Ā–Ķ —Ā–ĺ—Ā—ā–į–≤–Ķ–Ĺ–ł –ĺ–ī –Ņ–ĺ—Ā–Ķ–Ī–Ĺ–ł –Ĺ–ł–∑–ł –Ĺ–į –Ē–Ě–ö –ļ–ĺ–ł —Ā–Ķ –Ņ—Ä–Ķ–Ņ–ĺ–∑–Ĺ–į–≤–į–į—ā –Ņ–ĺ —Ä–Ķ–Ņ–Ľ–ł–ļ–į—Ü–ł—Ā–ļ–ł—ā–Ķ –∑–į—á–Ķ—ā–Ĺ–ł—á–ļ–ł –Ī–Ķ–Ľ–ļ–ĺ–≤–ł–Ĺ–ł, –ļ–ĺ–ł —Ā–Ķ –≤—Ä–∑—É–≤–į–į—ā –∑–į –ł–∑–≤–ĺ—Ä–Ĺ–ł—ā–Ķ –Ĺ–ł–∑–ł –∑–į –ī–į –Ņ–ĺ–ľ–ĺ–≥–Ĺ–į—ā –≤–ĺ –Ņ–ĺ–ī—Ä–Ķ–ī—É–≤–į—ö–Ķ—ā–ĺ –Ĺ–į –ī—Ä—É–≥–ł—ā–Ķ –Ī–Ķ–Ľ–ļ–ĺ–≤–ł–Ĺ–ł –Ņ–ĺ—ā—Ä–Ķ–Ī–Ĺ–ł –∑–į –Ē–Ě–ö-—Ä–Ķ–Ņ–Ľ–ł–ļ–į—Ü–ł—ė–į –Ĺ–į –ĺ–≤–ł–Ķ –ľ–Ķ—Ā—ā–į. –ē—É–ļ–į—Ä–ł–ĺ—ā—Ā–ļ–į—ā–į –∑–į—á–Ķ—ā–Ĺ–ł—á–ļ–į –Ī–Ķ–Ľ–ļ–ĺ–≤–ł–Ĺ–į –Ķ –ļ–ĺ–ľ–Ņ–Ľ–Ķ–ļ—Ā –ļ–ĺ—ė —Ā–Ķ —Ā–ĺ—Ā—ā–ĺ–ł –ĺ–ī —ą–Ķ—Ā—ā —Ä–į–∑–Ľ–ł—á–Ĺ–ł –Ņ–ĺ–ī–Ķ–ī–ł–Ĺ–ł—Ü–ł –ĺ–∑–Ĺ–į—á–Ķ–Ĺ–ł —Ā–ĺ –ł–ľ–Ķ—ā–ĺ –ł–∑–≤–ĺ—Ä–Ķ–Ĺ –Ņ—Ä–ł–∑–Ĺ–į–≤–į—á–ļ–ł –ļ–ĺ–ľ–Ņ–Ľ–Ķ–ļ—Ā (origin recognition complex –ł–Ľ–ł ORC). –Ď–į–ļ—ā–Ķ—Ä–ł—Ā–ļ–ł–ĺ—ā –∑–į—á–Ķ—ā–Ĺ–ł—á–ļ–ł –Ī–Ķ–Ľ–ļ–ĺ–≤–ł–Ĺ–į –Ķ –Ĺ–į—Ä–Ķ—á–Ķ–Ĺ –Ē–Ě–ö-–ź –Ī–Ķ–Ľ–ļ–ĺ–≤–ł–Ĺ–į. –í—Ä–Ķ–ľ–Ķ—ā—Ä–į–Ķ—ö–Ķ—ā–ĺ –Ĺ–į —Ä–Ķ–Ņ–Ľ–ł–ļ–į—Ü–ł—ė–į—ā–į –Ķ –∑–į–≤–ł—Ā–Ĺ–ĺ –ĺ–ī –Ņ–ĺ–ī—Ä–Ķ–ī—É–≤–į—ö–Ķ—ā–ĺ –Ĺ–į –ļ–ĺ–ľ–Ņ–Ľ–Ķ–ļ—Ā–ł—ā–Ķ –Ĺ–į –ł–∑–≤–ĺ—Ä–Ĺ–ł—ā–Ķ –ľ–Ķ—Ā—ā–į.
–ě–ī–ī–Ķ–Ľ–Ĺ–ł—ā–Ķ –Ķ—ā–į–Ņ–ł –ĺ–ī –Ņ–ĺ—á–Ķ—ā–Ĺ–į—ā–į —Ą–į–∑–į –Ĺ–į —Ä–Ķ–Ņ–Ľ–ł–ļ–į—Ü–ł—ė–į—ā–į —Ā–Ķ –Ņ–ĺ–ī–ĺ–Ī—Ä–ĺ –Ņ—Ä–ĺ—É—á–Ķ–Ĺ–ł –ļ–į—ė –Ī–į–ļ—ā–Ķ—Ä–ł–ł—ā–Ķ –ĺ—ā–ļ–ĺ–Ľ–ļ—É –ļ–į—ė –Ķ—É–ļ–į—Ä–ł–ĺ—ā–ł—ā–Ķ, –Ĺ–ĺ –Ĺ–Ķ–ļ–ĺ–Ľ–ļ—É –ļ–Ľ—É—á–Ĺ–ł –Ķ—ā–į–Ņ–ł —Ā–Ķ –∑–į–Ķ–ī–Ĺ–ł—á–ļ–ł –∑–į –ī–≤–Ķ—ā–Ķ –≥—Ä—É–Ņ–ł. –ü—Ä–≤–į—ā–į –Ķ—ā–į–Ņ–į –Ķ –Ņ—Ä–ĺ–ľ–Ķ–Ĺ–į –≤–ĺ –ļ–ĺ–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł—ė–į—ā–į –Ĺ–į –∑–į—á–Ķ—ā–Ĺ–ł—á–ļ–į—ā–į –Ī–Ķ–Ľ–ļ–ĺ–≤–ł–Ĺ–į, —ą—ā–ĺ –Ņ—Ä–Ķ–ī–ł–∑–≤–ł–ļ—É–≤–į –ĺ–≥—Ä–į–Ĺ–ł—á–Ķ–Ĺ–ĺ ‚Äě—ā–ĺ–Ņ–Ķ—ö–Ķ‚Äú (—ā–į–ļ–į —Ā–Ķ –Ĺ–į—Ä–Ķ–ļ—É–≤–į –ĺ–ī–ī–Ķ–Ľ—É–≤–į—ö–Ķ—ā–ĺ –Ĺ–į –ī–≤–Ķ—ā–Ķ –≤–Ķ—Ä–ł–≥–ł) –Ĺ–į –ī–≤–ĺ—Ā–Ņ–ł—Ä–į–Ľ–Ĺ–į—ā–į –Ē–Ě–ö –≤–Ķ–ī–Ĺ–į—ą –ī–ĺ –≤—Ä–∑—É–≤–į—á–ļ–ĺ—ā–ĺ –ľ–Ķ—Ā—ā–ĺ –∑–į –∑–į—á–Ķ—ā–Ĺ–ł—á–ļ–ł–ĺ—ā –Ī–Ķ–Ľ–ļ–ĺ–≤–ł–Ĺ–į. –ü–ĺ –ĺ–≤–į, –Ĺ–į –ļ–ĺ–ľ–Ņ–Ľ–Ķ–ļ—Ā–ĺ—ā –Ē–Ě–ö –Ľ–į–Ĺ–Ķ—Ü-–∑–į—á–Ķ—ā–Ĺ–ł–ļ —Ā–Ķ –Ņ—Ä–ł–ļ–Ľ—É—á—É–≤–į–į—ā —É—ą—ā–Ķ –ī–≤–Ķ –Ī–Ķ–Ľ–ļ–ĺ–≤–ł–Ĺ–ł, –ł —ā–ĺ–į –Ē–Ě–ö-—Ö–Ķ–Ľ–ł–ļ–į–∑–į –ł –Ē–Ě–ö-–Ņ—Ä–ł–ľ–į–∑–į. –ü–ĺ—á–Ķ—ā–Ĺ–į—ā–į –Ķ—ā–į–Ņ–į –Ĺ–į —Ä–Ķ–Ņ–Ľ–ł–ļ–į—Ü–ł—ė–į—ā–į –∑–į–Ņ–ĺ—á–Ĺ—É–≤–į —Ā–ĺ –į–ļ—ā–ł–≤–į—Ü–ł—ė–į –Ĺ–į —Ö–Ķ–Ľ–ł–ļ–į–∑–į—ā–į –ł –Ņ—Ä–ł–ľ–į–∑–į—ā–į, –ļ–į–ļ–ĺ –ł —Ā–ĺ –Ņ–ĺ–Ĺ–į—ā–į–ľ–ĺ—ą–Ĺ–ĺ—ā–ĺ –≤–ļ–Ľ—É—á—É–≤–į—ö–Ķ –Ĺ–į –Ē–Ě–ö-–Ņ–ĺ–Ľ–ł–ľ–Ķ—Ä–į–∑–į—ā–į. –ö–į—ė –Ņ—Ä–ĺ–ļ–į—Ä–ł–ĺ—ā–ł—ā–Ķ –Ņ–ĺ—Ā—ā–ĺ–ł –ī—Ä—É–≥–į —Ą–ĺ—Ä–ľ–į –Ĺ–į –ĺ–≤–ĺ—ė –Ķ–Ĺ–∑–ł–ľ - –Ē–Ě–ö-–Ņ–ĺ–Ľ–ł–ľ–Ķ—Ä–į–∑–į III. –í–ļ–Ľ—É—á–Ķ–Ĺ–ł —Ā–Ķ –ł –ī—Ä—É–≥–ł –Ī–Ķ–Ľ–ļ–ĺ–≤–ł–Ĺ–ł (–≤–ł–ī–ł –Ņ–ĺ–Ĺ–į—ā–į–ľ—É).
–†–Ķ–Ņ–Ľ–ł–ļ–į—Ü–ł—Ā–ļ–į –≤–ł—ô—É—ą–ļ–į
–ě–ī–ī–Ķ–Ľ—É–≤–į—ö–Ķ—ā–ĺ –Ĺ–į –ī–≤–Ķ—ā–Ķ —ą–į–Ī–Ľ–ĺ–Ĺ—Ā–ļ–ł –≤–Ķ—Ä–ł–≥–ł –ł —Ā–ł–Ĺ—ā–Ķ–∑–į—ā–į –Ĺ–į –Ĺ–ĺ–≤–ł —ú–Ķ—Ä–ļ–ł–Ĺ–ł –Ē–Ě–ö-–ľ–ĺ–Ľ–Ķ–ļ—É–Ľ–ł —Ā–ĺ–∑–ī–į–≤–į –Ņ–ĺ–ī–≤–ł–∂–Ĺ–į —Ä–Ķ–Ņ–Ľ–ł–ļ–į—Ü–ł—Ā–ļ–į –≤–ł—ô—É—ą–ļ–į. –í–ĺ –Ĺ–Ķ–į, –ī–≤–ĺ—Ā–Ņ–ł—Ä–į–Ľ–Ĺ–į—ā–į –Ē–Ě–ö –Ņ–ĺ—Ā—ā–ĺ—ė–į–Ĺ–ĺ —Ā–Ķ –ĺ–ī–ľ–ĺ—ā—É–≤–į –ł —Ā–Ķ –ļ–ĺ–Ņ–ł—Ä–į. –ě–ī–ľ–ĺ—ā—É–≤–į—ö–Ķ—ā–ĺ –Ĺ–į –Ē–Ě–ö —Ā–ĺ–∑–ī–į–≤–į –Ņ–ĺ—Ā–Ķ–Ī–Ĺ–ł –Ņ—Ä–ĺ–Ī–Ľ–Ķ–ľ–ł –∑–į –ļ–ĺ–ł –ľ–ĺ–∂–Ķ –ī–į —Ā–Ķ –Ĺ–į–Ņ—Ä–į–≤–ł –į–Ĺ–į–Ľ–ĺ–≥–ł—ė–į. –Ě–į –Ņ—Ä–ł–ľ–Ķ—Ä, –∑–į —Ä–į—Ā–ļ–ł–Ĺ—É–≤–į—ö–Ķ –Ĺ–į –ī–≤–Ķ –∂–ł—Ü–ł –ļ–ĺ–ł —Ā–Ķ —Ü–≤—Ä—Ā—ā–ĺ –ĺ–Ī–≤–ł–Ķ–Ĺ–ł –ľ–Ķ—ď—É—Ā–Ķ–Ī–Ĺ–ĺ –Ņ–ĺ—ā—Ä–Ķ–Ī–Ĺ–ĺ –Ķ –Ķ–Ĺ–Ķ—Ä–≥–ł—ė–į. –í–Ķ—ú–Ķ –ĺ–ī–ī–Ķ–Ľ–Ķ–Ĺ–ł—ā–Ķ –∂–ł—Ü–ł —Ā–Ķ —Ā—ā—Ä–Ķ–ľ–į—ā –ļ–ĺ–Ĺ –Ņ–ĺ–≤—ā–ĺ—Ä–Ĺ–ĺ –∑–į–ľ–ĺ—ā—É–≤–į—ö–Ķ, –į –ľ–Ķ—Ā—ā–ĺ—ā–ĺ –Ĺ–į –ļ–ĺ–Ķ –Ĺ–į—Ā—ā–į–Ĺ–į–Ľ–ĺ —Ä–į—Ā–ļ–ł–Ĺ—É–≤–į—ö–Ķ—ā–ĺ —Ā—ā–į–Ĺ—É–≤–į —É—ą—ā–Ķ –Ņ–ĺ—Ā–≤–ł–Ķ–Ĺ–ĺ.
–Ď–Ķ–Ľ–ļ–ĺ–≤–ł–Ĺ–ł—ā–Ķ –ĺ–ī —Ä–Ķ–Ņ–Ľ–ł–ļ–į—Ü–ł—Ā–ļ–į—ā–į –≤–ł—ô—É—ą–ļ–į —Ā–Ķ –Ĺ–į–ľ–Ķ–Ĺ–Ķ—ā–ł –∑–į —Ä–Ķ—ą–į–≤–į—ö–Ķ –Ĺ–į –≥–ĺ—Ä–Ķ—Ā–Ņ–ĺ–ľ–Ķ–Ĺ–į—ā–ł—ā–Ķ –Ņ—Ä–ĺ–Ī–Ľ–Ķ–ľ–ł. –Ē–Ě–ö-–Ņ–ĺ–Ľ–ł–ľ–Ķ—Ä–į–∑–ł—ā–Ķ –Ĺ–Ķ —Ā–Ķ —Ā–Ņ–ĺ—Ā–ĺ–Ī–Ĺ–ł –ī–į —ė–į –ĺ–ī–ľ–ĺ—ā–į–į—ā –ī–≤–ĺ—Ā–Ņ–ł—Ä–į–Ľ–Ĺ–į—ā–į –Ē–Ě–ö-–ľ–ĺ–Ľ–Ķ–ļ—É–Ľ–į, –∑–į —ą—ā–ĺ –Ķ –Ņ–ĺ—ā—Ä–Ķ–Ī–Ĺ–ĺ –Ķ–Ĺ–Ķ—Ä–≥–ł—ė–į –∑–į –ī–į —Ā–Ķ —Ä–į—Ā–ļ–ł–Ĺ–į—ā –≤–ĺ–ī–ĺ—Ä–ĺ–ī–Ĺ–ł—ā–Ķ –≤—Ä—Ā–ļ–ł –Ņ–ĺ–ľ–Ķ—ď—É –Ī–į–∑–Ĺ–ł—ā–Ķ –Ņ–į—Ä–ĺ–≤–ł –ļ–ĺ–ł –≥–ł –ī—Ä–∂–į—ā –≤–Ķ—Ä–ł–≥–ł—ā–Ķ –∑–į–Ķ–ī–Ĺ–ĺ. –ě–≤–į–į –∑–į–ī–į—á–į —ė–į –ĺ–Ī–į–≤—É–≤–į –Ē–Ě–ö-—Ö–Ķ–Ľ–ł–ļ–į–∑–į—ā–į, –ļ–ĺ—ė–į —ė–į –ļ–ĺ—Ä–ł—Ā—ā–ł –Ķ–Ĺ–Ķ—Ä–≥–ł—ė–į—ā–į –ĺ–ī –ź–Ę–ü –∑–į –ī–į —ė–į –ĺ–ī–ľ–ĺ—ā–į —ą–į–Ī–Ľ–ĺ–Ĺ—Ā–ļ–į—ā–į –Ē–Ě–ö. –ü–ĺ—ā–ĺ–į –Ķ–ī–ł–Ĺ–Ķ—á–Ĺ–ł—ā–Ķ –≤–Ķ—Ä–ł–≥–ł —Ā–Ķ –≤—Ä–∑—É–≤–į–į—ā —Ā–ĺ –≤—Ä–∑—É–≤–į—á–ļ–į –Ī–Ķ–Ľ–ļ–ĺ–≤–ł–Ĺ–į –∑–į –Ķ–ī–ł–Ĺ–Ķ—á–Ĺ–į –≤–Ķ—Ä–ł–≥–į (–ĺ–∑–Ĺ–į—á–Ķ–Ĺ –ļ–į–ļ–ĺ SSB, single-strand binding protein –ļ–į—ė –Ī–į–ļ—ā–Ķ—Ä–ł–ł—ā–Ķ –ł RPA –ļ–į—ė –Ķ—É–ļ–į—Ä–ł–ĺ—ā–ł—ā–Ķ). –ě–≤–į–į –Ī–Ķ–Ľ–ļ–ĺ–≤–ł–Ĺ–į –≥–ĺ —Ā–Ņ—Ä–Ķ—á—É–≤–į –Ņ–ĺ–≤—ā–ĺ—Ä–Ĺ–ĺ—ā–ĺ —Ā–Ņ–ł—Ä–į–Ľ–ł–∑–ł—Ä–į—ö–Ķ –Ĺ–į –≤–Ķ—Ä–ł–≥–ł—ā–Ķ. –ě–ī–ľ–ĺ—ā–į–Ĺ–į—ā–į –Ē–Ě–ö –ļ–ĺ—ė–į —Ā–Ķ —Ā–ĺ–∑–ī–į–≤–į –Ņ–ĺ —Ä–Ķ–Ņ–Ľ–ł–ļ–į—Ü–ł—Ā–ļ–į—ā–į –≤–ł—ô—É—ą–ļ–į —Ā–Ķ —Ā—ā—Ä–Ķ–ľ–ł –ī–į —Ä–ĺ—ā–ł—Ä–į –ł –ī–į —Ā–Ķ —Ā–≤–ł–≤–į. –ó–į –ī–į —Ā–Ķ —Ā–Ņ—Ä–Ķ—á–ł –ĺ–≤–į, –Ņ–ĺ—Ā—ā–ĺ–ł –Ķ–ī–Ķ–Ĺ –Ķ–Ĺ–∑–ł–ľ –ĺ–∑–Ĺ–į—á–Ķ–Ĺ –ļ–į–ļ–ĺ –Ē–Ě–ö-–≥–ł—Ä–į–∑–į (–ļ–į—ė –Ī–į–ļ—ā–Ķ—Ä–ł–ł—ā–Ķ) –ł–Ľ–ł —ā–ĺ–Ņ–ĺ–ł–∑–ĺ–ľ–Ķ—Ä–į–∑–į (–ļ–į—ė –Ķ—É–ļ–į—Ä–ł–ĺ—ā–ł—ā–Ķ), –ļ–ĺ—ė —ė–į –ł—Ā–Ņ—Ä–į–≤—É–≤–į –ł –ĺ–ī—Ä–∂—É–≤–į –≤–Ķ—Ä–ł–≥–į—ā–į.
 –ú–Ĺ–ĺ–≥—É –Ķ–Ĺ–∑–ł–ľ–ł —Ā–Ķ –≤–ļ–Ľ—É—á–Ķ–Ĺ–ł –≤–ĺ —Ä–Ķ–Ņ–Ľ–ł–ļ–į—Ü–ł—Ā–ļ–į—ā–į –≤–ł—ô—É—ą–ļ–į: 1. –Ĺ–Ķ–≤–ĺ–ī–Ķ—á–ļ–į (—Ā–Ľ–Ķ–ī–Ķ—á–ļ–į) –Ĺ–ł—ą–ļ–į; 2. –≤–ĺ–ī–Ķ—á–ļ–į –Ĺ–ł—ą–ļ–į; 3. –Ē–Ě–ö-–Ņ–ĺ–Ľ–ł–ľ–Ķ—Ä–į–∑–į; 4. –Ē–Ě–ö-–Ľ–ł–≥–į–∑–į; 5. –†–Ě–ö –Ņ—Ä–į—ė–ľ–Ķ—Ä; 6. –Ē–Ě–ö –Ņ—Ä–ł–ľ–į–∑–į; 7. –ě–ļ–į–∑–į–ļ–ł–Ķ–≤ —Ą—Ä–į–≥–ľ–Ķ–Ĺ—ā; 8. –Ē–Ě–ö-–Ņ–ĺ–Ľ–ł–ľ–Ķ—Ä–į–∑–į; 9. —Ö–Ķ–Ľ–ł–ļ–į–∑–į; 10. SSB –Ī–Ķ–Ľ–ļ–ĺ–≤–ł–Ĺ–į; 11. —ā–ĺ–Ņ–ĺ–ł–∑–ĺ–ľ–Ķ—Ä–į–∑–į.
–ú–Ĺ–ĺ–≥—É –Ķ–Ĺ–∑–ł–ľ–ł —Ā–Ķ –≤–ļ–Ľ—É—á–Ķ–Ĺ–ł –≤–ĺ —Ä–Ķ–Ņ–Ľ–ł–ļ–į—Ü–ł—Ā–ļ–į—ā–į –≤–ł—ô—É—ą–ļ–į: 1. –Ĺ–Ķ–≤–ĺ–ī–Ķ—á–ļ–į (—Ā–Ľ–Ķ–ī–Ķ—á–ļ–į) –Ĺ–ł—ą–ļ–į; 2. –≤–ĺ–ī–Ķ—á–ļ–į –Ĺ–ł—ą–ļ–į; 3. –Ē–Ě–ö-–Ņ–ĺ–Ľ–ł–ľ–Ķ—Ä–į–∑–į; 4. –Ē–Ě–ö-–Ľ–ł–≥–į–∑–į; 5. –†–Ě–ö –Ņ—Ä–į—ė–ľ–Ķ—Ä; 6. –Ē–Ě–ö –Ņ—Ä–ł–ľ–į–∑–į; 7. –ě–ļ–į–∑–į–ļ–ł–Ķ–≤ —Ą—Ä–į–≥–ľ–Ķ–Ĺ—ā; 8. –Ē–Ě–ö-–Ņ–ĺ–Ľ–ł–ľ–Ķ—Ä–į–∑–į; 9. —Ö–Ķ–Ľ–ł–ļ–į–∑–į; 10. SSB –Ī–Ķ–Ľ–ļ–ĺ–≤–ł–Ĺ–į; 11. —ā–ĺ–Ņ–ĺ–ł–∑–ĺ–ľ–Ķ—Ä–į–∑–į.
–í–ĺ–ī–Ķ—á–ļ–ł –Ĺ–ł–∑–ł –ł –Ĺ–Ķ–≤–ĺ–ī–Ķ—á–ļ–ł –Ĺ–ł–∑–ł
–ö–ĺ–ĺ—Ä–ī–ł–Ĺ–ł—Ä–į–Ĺ–į—ā–į —Ā–ł–Ĺ—ā–Ķ–∑–į –Ĺ–į –ī–≤–Ķ —ú–Ķ—Ä–ļ–ł–Ĺ–ł –Ĺ–ł–∑–ł –∑–į–ī–į–≤–į –≤–į–∂–Ķ–Ĺ –Ņ—Ä–ĺ–Ī–Ľ–Ķ–ľ –Ņ—Ä–ł –Ē–Ě–ö-—Ä–Ķ–Ņ–Ľ–ł–ļ–į—Ü–ł—ė–į—ā–į. –Ē–≤–Ķ—ā–Ķ –ľ–į—ė—á–ł–Ĺ–ł (—Ä–ĺ–ī–ł—ā–Ķ–Ľ—Ā–ļ–ł –ł–Ľ–ł –Ņ–į—Ä–Ķ–Ĺ—ā–į–Ľ–Ĺ–ł) –Ĺ–ł–∑–ł –ĺ–ī –Ē–Ě–ö —Ā–Ķ –Ņ—Ä–ĺ—ā–Ķ–≥–į–į—ā –≤–ĺ —Ā–Ņ—Ä–ĺ—ā–ł–≤–Ĺ–ł –Ĺ–į—Ā–ĺ–ļ–ł, –Ķ–ī–Ĺ–į—ā–į –ĺ–ī 5‚Ä≤ –ī–ĺ 3‚Ä≤ –ļ—Ä–į—ė–ĺ—ā, –į –ī—Ä—É–≥–į—ā–į –ĺ–ī 3‚Ä≤ –ī–ĺ 5‚Ä≤ –ļ—Ä–į—ė–ĺ—ā. –ú–Ķ—ď—É—ā–ĺ–į, —Ā–ł—ā–Ķ –Ņ–ĺ–∑–Ĺ–į—ā–ł –Ē–Ě–ö-–Ņ–ĺ–Ľ–ł–ľ–Ķ—Ä–į–∑–ł —ė–į –ļ–į—ā–į–Ľ–ł–∑–ł—Ä–į–į—ā —Ā–ł–Ĺ—ā–Ķ–∑–į—ā–į –Ĺ–į –Ē–Ě–ö —Ā–į–ľ–ĺ –≤–ĺ –Ķ–ī–Ķ–Ĺ –Ņ—Ä–į–≤–Ķ—Ü - –ĺ–ī 5‚Ä≤ –ļ–ĺ–Ĺ 3‚Ä≤ –ļ—Ä–į—ė–ĺ—ā, –ī–ĺ–ī–į–≤–į—ė—ú–ł –Ĺ—É–ļ–Ľ–Ķ–ĺ—ā–ł–ī–ł —Ā–į–ľ–ĺ –Ĺ–į 3‚Ä≤ –ļ—Ä–į—ė–ĺ—ā –Ĺ–į —Ä–į—Ā—ā–Ķ—á–ļ–į—ā–į –≤–Ķ—Ä–ł–≥–į. –Ē–ĺ–ļ–ĺ–Ľ–ļ—É —ú–Ķ—Ä–ļ–ł–Ĺ–ł—ā–Ķ –≤–Ķ—Ä–ł–≥–ł —Ā–Ķ —Ā–ł–Ĺ—ā–Ķ—ā–ł–∑–ł—Ä–į–į—ā –ļ–ĺ–Ĺ—ā–ł–Ĺ—É–ł—Ä–į–Ĺ–ĺ (–Ī–Ķ–∑ –Ņ—Ä–Ķ—Ā—ā–į–Ĺ), —ā–ĺ–≥–į—ą —ā–ł–Ķ –Ī–ł –ľ–ĺ—Ä–į–Ľ–Ķ –ī–į —Ā–Ķ —Ā–ł–Ĺ—ā–Ķ—ā–ł–∑–ł—Ä–į–į—ā –≤–ĺ —Ā–Ņ—Ä–ĺ—ā–ł–≤–Ķ–Ĺ –Ņ—Ä–į–≤–Ķ—Ü, –Ĺ–ĺ –ĺ–≤–į –Ĺ–Ķ —Ā–Ķ —Ā–Ľ—É—á—É–≤–į.
–†–Ķ—ą–Ķ–Ĺ–ł–Ķ—ā–ĺ –Ĺ–į –ĺ–≤–ĺ—ė –Ņ—Ä–ĺ–Ī–Ľ–Ķ–ľ –Ķ –≤–ĺ —ā–ĺ–į —ą—ā–ĺ —Ā–Ķ —Ā–ĺ–≥–Ľ–Ķ–ī–į–Ľ–ĺ –ī–Ķ–ļ–į —Ā–į–ľ–ĺ –Ķ–ī–Ĺ–į –ĺ–ī –ī–≤–Ķ—ā–Ķ —ú–Ķ—Ä–ļ–ł–Ĺ–ł –≤–Ķ—Ä–ł–≥–ł, –ĺ–∑–Ĺ–į—á–Ķ–Ĺ–į –ļ–į–ļ–ĺ –≤–ĺ–ī–Ķ—á–ļ–į –Ĺ–ł–∑–į, —Ā–Ķ —Ā–ł–Ĺ—ā–Ķ—ā–ł–∑–ł—Ä–į –Ĺ–Ķ–Ņ—Ä–Ķ—Ā—ā–į—ė–Ĺ–ĺ –≤–ĺ –Ņ—Ä–į–≤–Ķ—Ü –Ĺ–į –ī–≤–ł–∂–Ķ—ö–Ķ—ā–ĺ –Ĺ–į —Ä–Ķ–Ņ–Ľ–ł–ļ–į—Ü–ł—Ā–ļ–į—ā–į –≤–ł—ô—É—ą–ļ–į (–ĺ–ī 5‚Ä≤ –ļ–ĺ–Ĺ 3‚Ä≤ –ļ—Ä–į—ė–ĺ—ā). –í—ā–ĺ—Ä–į—ā–į —ú–Ķ—Ä–ļ–ł–Ĺ–į –≤–Ķ—Ä–ł–≥–į, –ĺ–∑–Ĺ–į—á–Ķ–Ĺ–į –ļ–į–ļ–ĺ –Ĺ–Ķ–≤–ĺ–ī–Ķ—á–ļ–į (—Ā–Ľ–Ķ–ī–Ķ—á–ļ–į) –Ĺ–ł—ą–ļ–į, —Ā–Ķ —Ā–ĺ–∑–ī–į–≤–į –ī–ł—Ā–ļ–ĺ–Ĺ—ā–ł–Ĺ—É–ł—Ä–į–Ĺ–ĺ –≤–ĺ –ľ–į–Ľ–ł —Ā–Ķ–≥–ľ–Ķ–Ĺ—ā–ł, –Ĺ–į—Ä–Ķ—á–Ķ–Ĺ–ł –ě–ļ–į–∑–į–ļ–ł–Ķ–≤–ł —Ą—Ä–į–≥–ľ–Ķ–Ĺ—ā–ł. –°–Ķ–ļ–ĺ—ė –ě–ļ–į–∑–į–ļ–ł–Ķ–≤ —Ą—Ä–į–≥–ľ–Ķ–Ĺ—ā —Ā–Ķ —Ā–ĺ–∑–ī–į–≤–į –≤–ĺ –Ņ—Ä–į–≤–Ķ—Ü–ĺ—ā 5‚Ä≤-3‚Ä≤ –ĺ–ī —Ā—ā—Ä–į–Ĺ–į –Ĺ–į –Ķ–Ĺ–∑–ł–ľ–ĺ—ā –Ē–Ě–ö-–Ņ–ĺ–Ľ–ł–ľ–Ķ—Ä–į–∑–į, —á–ł—ė –Ņ—Ä–į–≤–Ķ—Ü –Ĺ–į —Ā–ł–Ĺ—ā–Ķ–∑–į –Ķ –ĺ–Ī—Ä–į—ā–Ķ–Ĺ –≤–ĺ —Ā–Ņ–ĺ—Ä–Ķ–ī–Ī–į —Ā–ĺ —Ü–Ķ–Ľ–ĺ—Ā–Ĺ–ĺ—ā–ĺ –ī–≤–ł–∂–Ķ—ö–Ķ –Ĺ–į —Ä–Ķ–Ņ–Ľ–ł–ļ–į—Ü–ł—Ā–ļ–į—ā–į –≤–ł—ô—É—ą–ļ–į. –ü–ĺ—ā–ĺ–į, –ĺ–≤–ł–Ķ —Ą—Ä–į–≥–ľ–Ķ–Ĺ—ā–ł —Ā–Ķ —Ā–Ņ–ĺ—ė—É–≤–į–į—ā —Ā–ĺ –Ņ–ĺ–ľ–ĺ—ą –Ĺ–į –Ķ–Ĺ–∑–ł–ľ–ĺ—ā –Ĺ–į—Ä–Ķ—á–Ķ–Ĺ –Ē–Ě–ö-–Ľ–ł–≥–į–∑–į.
–Ě–į–ī–≤–ĺ—Ä–Ķ—ą–Ĺ–ł –≤—Ä—Ā–ļ–ł


