JeéûÀbi popelavûÙ báhem migrace pées ûrûÀn; ptûÀci v dolnûÙ áûÀsti fotografie udréƒujûÙ formaci ve tvaru pûÙsmene V, kterûÀ zefektivéuje letPéûÙklady tahovû§ch cest nákterû§ch dûÀlkovû§ch migranté₤
Migrace ptûÀké₤ je sezû°nnûÙ tah ptactva mezi jejich hnûÙzdiéÀti a zimoviéÀti. Na migraánûÙ cestu se pravidelná vydûÀvûÀ mûˋná neéƒ 20 % z vûÙce neéƒ 10 tisûÙc ptaáûÙch druhé₤. Opeéenci vykonûÀvajûÙcûÙ migraci se oznaáujûÙ jako taéƒnûÙ ái stáhovavûÙ ptûÀci. Dé₤vodem migrace je hlavná dostupnost potravy, obecnáji vytáéƒenûÙ dvou ái vûÙce biotopé₤, jejichéƒ vhodnost pro éƒivot ptûÀké₤ se sezû°nná promáéuje. Migrace je v ptûÀcûÙch geneticky zafixovûÀna, sociûÀlnûÙ uáenûÙ hraje druhotnou, i kdyéƒ nákdy pomárná vû§znamnou roli (napé. velkûˋ sociûÀlnûÙ druhy jako áûÀpi a dravci zûÙskûÀvajûÙ detaily o migraánûÙ trase uáenûÙm). VátéÀina ptûÀké₤ migruje v noci, coéƒ s sebou nese éadu vû§hod jako je vyhnutûÙ se predûÀtoré₤m nebo niéƒéÀûÙ teploty a tedy i menéÀûÙ riziko dehydratace.
K migraci dochûÀzûÙ podûˋl tahovû§ch cest, kterûˋ nejáastáji smáéujûÙ ze severu na jih s ré₤zná velkû§mi odchylkami k zûÀpadu ái vû§chodu. Tahovûˋ cesty vátéÀinou vedou podûˋl horskû§ch pûÀsem, vodnûÙch toké₤ a daléÀûÙch vû§raznû§ch topografickû§ch liniûÙ. VodnûÙ ptûÀci tûÀhnou podûˋl éek a pobéeéƒûÙ, plachtûÙcûÙ druhy (napé. dravci) vyhledûÀvajûÙ pevninskûˋ oblasti bohatûˋ na termickûˋ proudy, kterûˋ jim usnadéujûÙ let a pomûÀhajûÙ tak snûÙéƒit vysokûˋ energetickûˋ nûÀroky tahu. Vedle topografie se na vzniku tahovû§ch cest podûÙlely i glaciûÀlnûÙ cykly nebo historickûˋ rozéÀûÙéenûÙ ptûÀké₤. Stáhovavûˋ ptûÀky lze rozliéÀit podle dûˋlky tahu na migranty na dlouhou a krûÀtkou vzdûÀlenost. Migranti na dlouhou vzdûÀlenost typicky péekraáujûÙ velkou geografickou péekûÀéƒku, podnebnûˋ pûÀsmo nebo kontinent (napé. béehouéÀi, rybûÀci) a áasto migrujûÙ i tisûÙce, v extrûˋmnûÙch péûÙpadech i desetitisûÙce kilometré₤ daleko. Migranti na krûÀtkou vzdûÀlenost migrujûÙ spûÙéÀe lokûÀlná (napé. kavka obecnûÀ, kterûÀ migruje ze stéednûÙ do jihozûÀpadnûÙ Evropy). éada ptûÀké₤ vykonûÀvûÀ vû§éÀkovou migraci, tedy péesun mezi ré₤znû§mi nadmoéskû§mi vû§éÀkami (typicky z vû§éÀe poloéƒenû§ch hnûÙzdiéÀéË do nûÙéƒe poloéƒenû§ch zimoviéÀéË). Nákdy nemigrujûÙ véÀichni jedinci, ale pouze áûÀst populace druhu ã napé. jen uráitû§ poddruh ái jen dospálûÙ jedinci. Takovûˋto péûÙpady se oznaáujûÙ jako áûÀsteánûÀ migrace.
NaáasovûÀnûÙ migrace se éûÙdûÙ geneticky fixovanû§m endogennûÙm (vniténûÙm) cirkanuûÀlnûÙm rytmem seéûÙzenû§m podle dûˋlky svátla báhem dne. Vedle toho majûÙ na naáasovûÀnûÙ vliv i genetickûÀ vû§bava, poáasûÙ, sociûÀlnûÙ stimuly, biologickûÀ zdatnost, teplota nebo dostupnost potravy. PtûÀci pro navigaci pouéƒûÙvajûÙ magnetickûˋ pole Zemá, olfaktorickûˋ (áichovûˋ) vnûÙmûÀnûÙ, vizuûÀlnûÙ prvky v terûˋnu i polohu Slunce a hvázd. Obáas se stane, éƒe se ptûÀci zatoulajûÙ, coéƒ je tradiáná péipisovûÀno na vrub navigaánûÙm chybûÀm jedincé₤, nicmûˋná novájéÀûÙ bûÀdûÀnûÙ ukazuje, éƒe zbloudánûÙ zûÀroveé umoéƒéuje ptûÀké₤m objevovat novûÀ hnûÙzdiéÀtá a zimoviéÀtá.
U stáhovavû§ch ptûÀké₤ se vyvinula éada fyziologickû§ch, hormonûÀlnûÙch i neurologickû§ch adaptacûÙ, kterûˋ jim pomûÀhajûÙ vyrovnat se s extrûˋmnûÙ povahou migrace. K támto adaptacûÙm patéûÙ schopnost tála navû§éÀit zûÀsoby tuku péed migracûÙ, kontrola spûÀnku, smréÀéËovûÀnûÙ orgûÀné₤ péed tahem nebo naáasovûÀnûÙ péepeéovûÀnûÙ. Se souáasnû§m globûÀlnûÙm oteplovûÀnûÙm probûÙhûÀ u stáhovavû§ch ptûÀké₤ éada velkû§ch zmán; mj. dochûÀzûÙ k áasnájéÀûÙm odleté₤m do zimoviéÀéË. Ochrana taéƒnû§ch druhé₤ je organizaáná nûÀroánûÀ a vyéƒaduje koordinaci mnoha stûÀté₤, proáeéƒ byla vytvoéena éada û¤mluv jako je tzv. BonnskûÀ û¤mluva, v rûÀmci kterûˋ byla sjednûÀna i Dohoda o ochraná africko-euroasijskû§ch stáhovavû§ch vodnûÙch ptûÀké₤.
Migrace ptûÀké₤ fascinovala lidstvo od nepamáti. O tahu ptûÀké₤ se objevujûÙ zmûÙnky uéƒ ve Starûˋm zûÀkoná ái Homûˋrová Illiadá. Taéƒnûˋ ptactvo studoval i staroéeckû§ klasik Aristotelûˋs, kterû§ vyslovil myéÀlenku, éƒe vlaéÀtovky zimujûÙ v bahná moáûÀlé₤. Tato hypotûˋza se stala péevlûÀdajûÙcûÙm vysvátlenûÙm kaéƒdoroánûÙho mizenûÙ vlaéÀtovek po 2 tisûÙce let a pré₤lom nastal aéƒ ve druhûˋ poloviná 19. stoletûÙ s nûÀstupem modernûÙch vádeckû§ch pokusé₤ a metod, péedevéÀûÙm krouéƒkovûÀnûÙ. To je dodnes nejáastájéÀûÙ metodou studia ptaáûÙ migrace. DaléÀûÙm milnûÙkem ve studiu taéƒnû§ch ptûÀké₤ nastal s nûÀstupem telemetrickû§ch zaéûÙzenûÙ, zejmûˋna ultralehkû§ch geolokûÀtoré₤ (GLS). V poloviná 10. let 21. stoletûÙ doéÀlo i k prvnûÙm testé₤m telemetrickû§ch zaéûÙzenûÙ o vûÀze kolem 1 g, kterûÀ lze péipevnit i na malûˋ druhy pávcé₤. I pées pomárná intenzivnûÙ zûÀjem vádcé₤ a pouéƒitûÙ modernûÙch technologiûÙ zé₤stûÀvûÀ éada otûÀzek okolo migrace ptûÀké₤ neobjasnána.
Charakteristika migrace
Z hlediska pohybu lze ptûÀky rozdálit na:
stûÀlûˋ (ptûÀci, kteéûÙ éƒijûÙ ve stejnûˋ oblasti po celû§ rok);
potulnûˋ (nákterûˋ populace ptûÀké₤, jako jsou nákterûˋ sû§kory ái pánkavy, kterûˋ se mimo hnûÙzdnûÙ obdobûÙ potulujûÙ po okolnûÙ krajiná bez specifickûˋho éûÀdu);
Tahovûˋ cesty áûÀpé₤ bûÙlû§ch vedou péevûÀéƒná mezi evropskû§mi hnûÙzdiéÀti (zelenou barvou) a africkû§mi zimoviéÀti (modée)
Z celkovûˋho poátu vûÙce neéƒ 10 000 druhé₤ ptûÀké₤ se za taéƒnûˋ povaéƒuje kolem 1850 druhé₤ áili necelû§ch 20 %.[2][3] Migraci ptûÀké₤ lze definovat jako kaéƒdoroánûÙ hromadnû§ péesun ptûÀké₤ mezi jejich hnûÙzdiéÀti a zimoviéÀti.[4] Pojem ãzimoviéÀtáã mé₤éƒe bû§t problematickû§, a to zvlûÀéÀtá u druhé₤ péekraáujûÙcûÙch rovnûÙk, takéƒe se nákdy mûÙsto vû§razé₤ ãhnûÙzdiéÀtáã a ãzimoviéÀtáã pouéƒûÙvajûÙ pojmy ãhnûÙzdicûÙã a ãnehnûÙzdicûÙã oblast.[5] PrimûÀrnûÙm dé₤vodem migrace je dostupnost potravy, resp. vytáéƒenûÙ dvou ái vûÙce biotopé₤, jejichéƒ vhodnost se pro éƒivot ptûÀké₤ sezû°nná promáéuje.[6] K támto prománûÀm patéûÙ vedle zmán v dostupnosti potravy i zmány habitatu, péûÙpadná poáasûÙ.[7] PoáasûÙ, resp. pokles teplot mé₤éƒe bû§t pro nákterûˋ druhy druhotnû§ motivujûÙcûÙ faktor migrace, nicmûˋná éada druhé₤ ã jako napé. nákteéûÙ kolibéûÙci ã dokûÀéƒe péeákat tuhûˋ zimy v hnûÙzdiéÀtûÙch, pokud majûÙ dostatek potravy.[4] Navzdory dlouhodobûˋmu zûÀjmu vádcé₤ je migraánûÙ chovûÀnûÙ ptûÀké₤ z velkûˋ áûÀsti stûÀle nedostateáná vysvátleno.[8]
Migrace ptûÀké₤ se odehrûÀvûÀ péevûÀéƒná na severnûÙ polokouli.[3] Dé₤vodem je, éƒe vátéÀina pevniny severnûÙ polokoule se nachûÀzûÙ v mûÙrnû§ch a subpolûÀrnûÙch zemápisnû§ch éÀûÙékûÀch, ve kterû§ch dochûÀzûÙ v zimá k nedostatku potravy, coéƒ zpé₤sobuje migraci ptûÀké₤ na jih, nákdy aéƒ na jiéƒnûÙ polokouli. Ponákud odliéÀnûÀ je situace u moéskû§ch ptûÀké₤, u kterû§ch je vátéÀûÙ pravdápodobnost migrace na jiéƒnûÙ polokouli. Dé₤vody jsou spojovûÀny s vátéÀûÙmi plochami oceûÀné₤ ái vátéÀûÙm mnoéƒstvûÙm vhodnû§ch ostrové₤ k hnûÙzdánûÙ.[9]
Migrace ptûÀké₤ neprobûÙhûÀ pouze ve vzduchu. VátéÀina tuáéûÀké₤ migruje ve vodá plavûÀnûÙm a v nákterû§ch péûÙpadech i ché₤zûÙ ã napé. tuáéûÀk cûÙsaéskû§ ujde z vody do kolonie na ledu i pées 100 km jednûÙm smárem.[10][11] Podobná pochoduje i tetéûÙvek douglaskovû§, kterû§ vykonûÀvûÀ kaéƒdoroánûÙ vû§éÀkovou migraci o nákolika stech vû§éÀkovû§ch metrech péevûÀéƒná pochodem po zemi.[12] Pravidelnû§ migraánûÙ pochod vykonûÀvûÀ i zûÀpadnûÙ populace australskûˋho báéƒceemua hnádûˋho.[13]
MigraánûÙ let
HoufovûÀnûÙ éÀpaáké₤ na Jutskûˋm poloostrová péed migraánûÙm letem veéÀlo ve znûÀmost dûÙky obrovskû§m hejné₤m éÀpaáké₤, kterûÀ za soumraku a za û¤svitu ãtanáûÙã po obloze. V DûÀnsku se támto masivnûÙm hejné₤m éûÙkûÀ sort sol (ãáernûˋ slunceã)
K migraci typicky dochûÀzûÙ ve vlnûÀch, tzn. v rûÀmci uráitûˋho druhu nemigrujûÙ véÀichni ptûÀci najednou, ale ve skupinûÀch. Doba odletu zûÀvisûÙ mj. na áasu, mûÙstnûÙch podmûÙnkûÀch a situaci v mûÙstá péûÙletu.[14] Je pomárná báéƒnûˋ, éƒe ré₤znûˋ vákovûˋ skupiny nebo rozdûÙlnûÀ pohlavûÙ majûÙ ré₤znûˋ migraánûÙ vzorce. NapéûÙklad samice slavûÙké₤ modrûÀáké₤ ve vû§chodnûÙ FennoskandinûÀvii[15] ái samice jespûÀké₤ obecnû§ch se vydûÀvajûÙ na migraánûÙ cestu déûÙve neéƒ samci.[16] U éady daléÀûÙch druhé₤ (napé. vodnûÙ ptûÀci vodouéÀ bahennûÙ, jespûÀk kéivozobû§, jespûÀk rezavû§, jespûÀk éÀedû§) migrujûÙ dospálci déûÙve neéƒ juvenilnûÙ jedinci.[17] Sû§korky rodé₤ Cyanistes a Parus migrujûÙ pouze v prvnûÙm roce éƒivota.[18] Obecná platûÙ, éƒe jarnûÙ migrace bû§vûÀ rychlejéÀûÙ neéƒ podzimnûÙ.[19]
ZatûÙmco nákterûˋ druhy migrujûÙ samostatná (napé. kolibéûÙci[20] a noánûÙ migranti), jinûÙ ptûÀci migrujûÙ v hejnu. ShromaéƒáovûÀnûÙ ptûÀké₤ do hejn se nazû§vûÀ houfovûÀnûÙ. Tah v hejnu mûÀ éadu vû§hod jako kolektivnûÙ rozhodovûÀnûÙ (ãvûÙc hlav vûÙc vûÙã), dûÙky kterûˋmu ptûÀci mohou snadnáji nalûˋzt vhodnûÀ mûÙsta k odpoáinku, efektivnáji sbûÙrat potravu, vybrat nejvhodnájéÀûÙ tahovou cestu nebo snadnáji a déûÙve odhalit predûÀtora. ZatûÙmco v nákterû§ch hejnech se ptûÀci dréƒûÙ daleko od sebe (napé. nákteéûÙ pávci udréƒujûÙ vzûÀjemnou vzdûÀlenost i 50 m a vûÙce), jinûˋ druhy se dréƒûÙ tak tásná u sebe, éƒe se takéka dotû§kajûÙ koneáky kéûÙdel (napé. husy nebo labutá).[21] CestovûÀnûÙ v hejnech pomûÀhûÀ sniéƒovat energetickûˋ nûÀroky letu.[22] Velikost hejn se mé₤éƒe razantná liéÀit podle druhu od nákolika jedincé₤ po tisûÙce ptûÀké₤. Hejna éÀpaáké₤ mohou zahrnovat i miliony jedincé₤.[23][24]
NákterûÀ hejna vátéÀûÙch druhé₤ ptûÀké₤, jako jsou pelikûÀni, jeéûÀbi, kormorûÀni, husy nebo labutá, tvoéûÙ v letu formaci ve tvaru pûÙsmene V. Tato formace umoéƒéuje jedincé₤m po stranûÀch snûÙéƒit odpor vzduchu, a tedy i mnoéƒstvûÙ energie nutnûˋ k letu. Odhaduje se, éƒe ptûÀci takto uéÀetéûÙ 20ã30 % energie oproti samostatnûˋmu letu.[25] Jedincé₤m v áele formace takovûÀ û¤spora neplyne, a proto se ptûÀci v áele áasto stéûÙdajûÙ. PtûÀci dokonce dovedou synchronizovat mûÀvûÀnûÙ kéûÙdel tak, aby co nejlûˋpe vyuéƒûÙvali vztlakovû§ch proudé₤ a naopak se efektivná vyhnuli sestupnû§m proudé₤m, kterûˋ zpé₤sobujûÙ mûÀvûÀnûÙm kéûÙdly ptûÀci péed nimi.[26] U jespûÀké₤ rezavû§ch bylo namáéeno, éƒe péi pohybu v hejnech lûˋtajûÙ o 5 km/h rychleji neéƒ péi samostatnûˋm letu.[27] U pelikûÀné₤ bûÙlû§ch bylo zaznamenûÀno, éƒe péi letu ve formaci ve tvaru V majûÙ pomalejéÀûÙ srdeánûÙ tep a nemusûÙ tolik mûÀvat kéûÙdly jako samostatná letûÙcûÙ jedinci.[28] DaléÀûÙ vû§hodou tûˋto formace je snadnûˋ udréƒovûÀnûÙ vizuûÀlnûÙho kontaktu ptûÀké₤ mezi sebou.[29]
MigraánûÙ let probûÙhûÀ v ré₤znû§ch nadmoéskû§ch vû§éÀkûÀch. V oblasti stéedoevropskû§ch nûÙéƒin ptûÀci tûÀhnou mezi 0,2ã1,5 km nad zemûÙ.[30] Obecná platûÙ, éƒe dlouhûˋ migraánûÙ lety probûÙhajûÙ ve vyéÀéÀûÙch nadmoéskû§ch vû§éÀkûÀch neéƒ krûÀtkûˋ migraánûÙ lety. VyéÀéÀûÙ nadmoéskûÀ vû§éÀka sice poskytuje vhodnájéÀûÙ podmûÙnky k letu (niéƒéÀûÙ teplota, silnájéÀûÙ vûÙtr), avéÀak vystoupat do takovûˋ vû§éÀky je energeticky nûÀroánûˋ. Proto se v dosaéƒenûˋ vû§éÀce ptûÀci dréƒûÙ co nejdeléÀûÙ dobu.[31]
K nejvátéÀûÙm ãvzduéÀnû§m horolezcé₤mã patéûÙ husy indickûˋ, kterûˋ pravidelná lûˋtajûÙ pées himûÀlajskûˋ pohoéûÙ. Husy se báéƒná vyskytujûÙ ve vû§éÀkûÀch kolem 5000ã6000 m n. m., avéÀak mohou lûˋtat i ve vû§éÀkûÀch péesahujûÙcûÙch 7000 m n. m.[32] Vû§éÀkovû§ rozdûÙl 6000 m mohou péekonat péibliéƒná báhem osmi hodin.[33] ZatûÙmco husy indickûˋ jsou do takovû§ch vû§éÀek péinuceny letát kvé₤li geografickûˋ péekûÀéƒce, béehouéÀi áernoocasûÙ stoupajûÙ do nákolikakilometrovû§ch vû§éÀek kvé₤li hledûÀnûÙ vhodnû§ch vátré₤ a aby se vyhnuli horkûˋmu vzduchu v niéƒéÀûÙch polohûÀch.[34] Druhy pohybujûÙcûÙ se pomocûÙ plachtánûÙ (napé. dravci) se vátéÀinou pohybujûÙ v nadmoéskû§ch vû§éÀkûÀch do 1500 m, kde je dostatek termickû§ch proudé₤.[35] Moée, kde tyto vzduéÀnûˋ proudy chybûÙ, péekonûÀvajûÙ plachtûÙcûÙ druhy nad û¤éƒinami (v Evropá patéûÙ k áasto vyuéƒûÙvanû§m péechodé₤m Gibraltarskû§ pré₤liv[36] nebo MessinskûÀ û¤éƒina).[37]
Tahovûˋ cesty
PodrobnájéÀûÙ informace naleznete v álûÀnku TahovûÀ cesta.
Mapa sváta zobrazujûÙcûÙ moéƒnûˋ rozdálenûÙ globûÀlnûÙch migraánûÙch cest ptûÀké₤ do nûÀsledujûÙcûÙch tahovû§ch cest:[5]
PacifickûÀ
MississippskûÀ
AtlantickûÀ
Vû§chodo-atlantickûÀ
áernomoésko-stéedozemnûÙ
ZûÀpadoasijsko-vû§chodoafrickûÀ
StéedoasijskûÀ
Vû§chodoasijsko-australasijskûÀ
PtûÀci se na migraánûÙ cestu vydûÀvajûÙ z ré₤znû§ch konáin, avéÀak báhem tahu se jejich pohyb postupná koncentruje do relativná ustûÀlenû§ch tras, jiméƒ se éûÙkûÀ tahovûˋ cesty (nákdy tûˋéƒ obecnáji migraánûÙ cesty ái migraánûÙ trasy).[38][39] Tahovûˋ cesty typicky vedou podûˋl horskû§ch pûÀsem, oceûÀnskûˋho pobéeéƒûÙ a daléÀûÙch geografickû§ch liniûÙ. VodnûÙ ptûÀci áasto migrujûÙ podûˋl éek a vodnûÙch toké₤, kde mohou nalûˋzt potravu. Dravci a daléÀûÙ plachtûÙcûÙ druhy vyhledûÀvajûÙ trasy bohatûˋ na stoupavûˋ vzduéÀnûˋ proudy a termûÀlnûÙ vátry, kterûˋ jim usnadéujûÙ let, a vyhû§bajûÙ se otevéenû§m vodnûÙm plochûÀm, kde se stoupavûˋ vzduéÀnûˋ proudy nevyskytujûÙ. Je pomárná báéƒnûˋ, éƒe ptûÀci volûÙ péi cestá do zimoviéÀtá jinou trasu neéƒ péi cestá zpát.[40]
Tahovûˋ cesty jen vzûÀcná péedstavujûÙ tu nejkratéÀûÙ moéƒnou variantu, jak se dostat z hnûÙzdiéÀtá do zimoviéÀtá. KlûÙáovûˋ faktory, kterûˋ ovlivéujûÙ dûˋlku cesty, jsou topografie, poáasûÙ a stanoviéÀtá bohatûÀ na potravu.[41] Nákdy ptûÀci nûÀsledujûÙcûÙ deléÀûÙ trasu dûÙky zdádánûÙ migraánûÙch gené₤ svû§ch péedké₤, kteéûÙ vedli migraci po uráitûˋ trase, kterûÀ véÀak jiéƒ nepéedstavuje tu nejvû§hodnájéÀûÙ nebo nejkratéÀûÙ moéƒnost. PéûÙkladem mé₤éƒe bû§t drozd zûÀpadnûÙ, malû§ severoamerickû§ pávec, jehoéƒ populace byla báhem poslednûÙ doby ledovûˋ rozdálena do dvou geneticky vû§razná odliéÀnû§ch skupin, z nichéƒ jedna zé₤stala ve vnitrozemûÙ a druhûÀ byla vytlaáena k aljaéÀskûˋmu pobéeéƒûÙ. Báhem migrace véÀak drozdi z AljaéÀky nejdéûÙve letûÙ tisûÙce kilometré₤ na vû§chod, neéƒ zamûÙéûÙ na jih smárem ke svû§m stéedoamerickû§m hnûÙzdiéÀtûÙm, mûÙsto aby nûÀsledovali kratéÀûÙ trasu podûˋl zûÀpadnûÙho pobéeéƒûÙ.[42] áasto se stûÀvûÀ, éƒe sousedûÙcûÙ populace tûˋhoéƒ druhu migrujûÙ do zimoviéÀéË jinû§mi trasami nebo migrujûÙ do zimoviéÀéË v jinû§ch lokalitûÀch. Oblasti mezi támito populacemi se éûÙkûÀ migraánûÙ rozhranûÙ. Tento jev zpé₤sobujûÙ geografickûˋ bariûˋry i historickûˋ glaciûÀlnûÙ cykly, kterûˋ formovaly vû§skyt a pohyb ptactva. MigraánûÙ rozhranûÙ se vyskytujûÙ i na û¤zemûÙ áeska ã zatûÙmco nákterûˋ moravskûˋ populace ptûÀké₤ tûÀhnou na jih jihovû§chodnûÙ tahovou cestou, ptûÀci ze zûÀpadnûÙ áûÀsti zemá mohou zvolit jihozûÀpadnûÙ migraánûÙ cestu.[43] PéûÙkladem mohou bû§t rûÀkosnûÙk obecnû§, rehek domûÀcûÙ[44] nebo áûÀp bûÙlû§.[45]
HoufovûÀnûÙ vlaéÀtovek obecnû§ch v Braniborsku péed odletem do Afriky
Nejáastáji dochûÀzûÙ k migraci ze severu na jih, s menéÀûÙ ái vátéÀûÙ odchylkou k vû§chodu ái zûÀpadu. Na severnûÙ polokouli ptûÀci typicky odlûˋtajûÙ na jaée do hnûÙzdiéÀéË v severnûÙch oblastech, kde zaáûÙnajûÙ kvûˋst rostliny a mnoéƒûÙ se hmyz, a na podzim se vracejûÙ do teplejéÀûÙch zimoviéÀéË na jihu. Na jiéƒnûÙ polokouli je proces obrûÀcenû§.[46] Vedle lepéÀûÙ vû§táéƒnosti zdrojé₤ je vû§hodou tahé₤ po ose severãjih oproti migracûÙm na trase vû§chodãzûÀpad lepéÀûÙ vyuéƒitûÙ dennûÙho svátla; zejmûˋna v polûÀrnûÙch a subpolûÀrnûÙch oblastech dlouhûˋ letnûÙ dny poskytujûÙ ptûÀké₤m vûÙce áasu na shûÀnánûÙ potravy a krmenûÙ mlûÀáat. DûÙky tomu mûÙvajûÙ taéƒnûÙ ptûÀci vátéÀûÙ sné₤éÀky neéƒ stûÀlûÙ ptûÀci.[47] Tahovûˋ cesty nákterû§ch druhé₤ vedou spûÙéÀe z vû§chodu na zûÀpad. PéûÙkladem mohou bû§t nákterûˋ africkûˋ druhy migrujûÙcûÙ mezi vyprahlû§mi lûˋty zûÀpadnûÙ Afriky a vlhkû§mi zimami na vû§chodá nebo nákterûˋ druhy moéskû§ch ptûÀké₤, vrubozubû§ch a bahéûÀké₤, kteéûÙ na jaée opouéÀtájûÙ zimoviéÀtá v arktickû§ch oblastech a nejdéûÙve se vydûÀvajûÙ vû§chodnûÙm ái zûÀpadnûÙm smárem podûˋl severnûÙch hranic kontinenté₤, neéƒ zamûÙéûÙ zahnûÙzdit do vnitrozemûÙ.[48] Obecná platûÙ, éƒe taéƒnûÙ ptûÀci hnûÙzdûÙcûÙ na severnûÙ polokouli majûÙ deléÀûÙ migraánûÙ trasy neéƒ ptûÀci hnûÙzdûÙcûÙ na jiéƒnûÙ polokouli.[49]
Koncept tahovû§ch cest je éÀiroce vyuéƒûÙvûÀn v managementu a ochraná ptaáûÙch populacûÙ, kterûˋ vyéƒadujûÙ mezinûÀrodnûÙ spoluprûÀci. V táchto péûÙpadech jsou tahovûˋ cesty znaáná zjednoduéÀenûˋ a nutná se ztrûÀcûÙ éada informacûÙ. NereflektujûÙ tah konkrûˋtnûÙch jedincé₤, ale spûÙéÀe globûÀlnûÙ trendy.[5]MezinûÀrodnûÙ svaz ochrany péûÙrody rozliéÀuje átyéi hlavnûÙ tahovûˋ cesty: mezi JiéƒnûÙ a SevernûÙ Amerikou, Afrikou a Evropou, stéednûÙ a jiéƒnûÙ AsiûÙ, vû§chodnûÙ AsiûÙ a AustralasiûÙ.[50]
VátéÀina stáhovavû§ch ptûÀké₤ báhem migrace zastavuje na vhodnû§ch stanoviéÀtûÙch. Tato stanoviéÀtá poskytujûÙ ptûÀké₤m potravu nutnou k doplnánûÙ energetickû§ch zûÀsob, bezpeánûˋ û¤toáiéÀtá péed predûÀtory a nákdy i ochranu péed extrûˋmnûÙm poáasûÙm jako jsou tropickûˋ bouée. Tahovûˋ zastûÀvky jsou pro ptûÀky áasto naprosto klûÙáovûˋ. NákteéûÙ taéƒnûÙ ptûÀci ztrûÀcûÙ v prvnûÙ áûÀsti migraánûÙho letu i polovinu svûˋ vûÀhy a bez potravnûÙch zastûÀvek, kde dojde k opátovnûˋmu nabrûÀnûÙ energie, by nebyli schopni tah dokonáit.[51] NákteéûÙ ptûÀci strûÀvûÙ aéƒ 80 % doby migrace na odpoáinkovû§ch stanoviéÀtûÙch. Doba strûÀvenûÀ na odpoáinkovûˋm stanoviéÀti je ovlivnána fyzickû§m stavem ptûÀké₤, jejich biorytmy (resp. endogennûÙmi hodinami) a environmentûÀlnûÙmi podmûÙnkami jako je poáasûÙ, mûÙra predace a dostupnost potravy.[52] TaéƒnûÙ ptûÀci vyskytujûÙcûÙ se na tahovû§ch zastûÀvkûÀch mezi hnûÙzdiéÀti a zimoviéÀti se v rûÀmci danûˋ lokality oznaáujûÙ jako protahujûÙcûÙ druhy (péûÙpadná pré₤taéƒnûÙ druhy).[53]
MigraánûÙ druhy ptûÀké₤ hrajûÙ dé₤leéƒitou roli v mûÙstnûÙch ekosystûˋmech. StáhovavûÙ ptûÀci pomûÀhajûÙ s opylovûÀnûÙm rostlin i roznûÀéÀenûÙm semen, kontrolujûÙ populaci hmyzu a slouéƒûÙ jako koéist mûÙstnûÙch predûÀtoré₤ ái naopak lovûÙ mûÙstnûÙ éƒivoáichy.[55]Jikry nákterû§ch druhé₤ ryb se dokonce mohou rozptylovat do vzdûÀlenû§ch a izolovanû§ch vodnûÙch ploch skrze ptaáûÙ trûÀvicûÙ trakt.[56] MasivnûÙ péesuny ptûÀké₤ z jednoho mûÙsta na druhûˋ mohou zlepéÀovat odolnost mûÙstnûÙch ekosystûˋmu nûÀsledkem propojenûÙ biotopé₤, coéƒ zvyéÀuje jejich druhovou rozmanitost.[57] Stáhovavûˋ druhy véÀak nemusûÙ péinûÀéÀet jen pozitiva; s ptaáûÙmi migranty péichûÀzûÙ i hrozba masivnûÙho éÀûÙéenûÙ nemocûÙ a viré₤.[58] Za péûÙklady mé₤éƒe slouéƒit rozéÀûÙéenûÙ neotropickû§ch klûÙéÀéËat ve Spojenû§ch stûÀtech nûÀsledkem jejich introdukce na tálech ptûÀké₤[59] nebo éÀûÙéenûÙ ptaáûÙ chéipky kolem sváta.[60]
MigraánûÙ strategie
Existuje celûÀ éada migraánûÙch strategiûÙ, kterûˋ se nejáastáji rozdálujûÙ podle dûˋlky (migrace na krûÀtkou vs. migrace na dlouhou vzdûÀlenost) nebo péevaéƒujûÙcûÙ dennûÙ doby migrace (dennûÙ vs. noánûÙ). Mezi migraánûÙmi strategiemi nenûÙ jasná stanovenûÀ hranice. NezéûÙdka se stûÀvûÀ, éƒe ré₤znûˋ populace ái jedinci v rûÀmci tûˋhoéƒ druhu praktikujûÙ odliéÀnûˋ migraánûÙ strategie (napé. áûÀst populace mé₤éƒe migrovat na dlouhou vzdûÀlenost, áûÀst na krûÀtkou vzdûÀlenost a áûÀst populace je stûÀlûÀ).[61][62]
Migrace na krûÀtkou vzdûÀlenost
Kavka obecnûÀ, typickû§ evropskû§ ptaáûÙ migrant na krûÀtkou vzdûÀlenost
Jako migrace na krûÀtkou vzdûÀlenost se oznaáujûÙ tahy ptûÀké₤ bez vû§znamnájéÀûÙch geografickû§ch péekûÀéƒek, typicky v rûÀmci kontinenté₤ nebo podnebnû§ch pûÀsé₤.[63][61] MaximûÀlnûÙ vzdûÀlenost, kterûÀ by odliéÀovala migraci na krûÀtkou vzdûÀlenosti od dlouhûˋ se vátéÀinou neuvûÀdûÙ nebo se liéÀûÙ podle zdroje. Napé. americkû§ ornitolog John H. Rappole povaéƒuje hornûÙ hranici vzdûÀlenosti tahu migranté₤ na krûÀtkou vzdûÀlenost kolem 2 tisûÙc kilometré₤; deléÀûÙ tahy povaéƒuje za migrace na dlouhou vzdûÀlenost.[63]
Migranti na krûÀtkou vzdûÀlenost jsou typicky druhy z tropickûˋho, subtropickûˋho a jiéƒnûÙch oblastûÙ mûÙrnûˋho podnebnûˋho pûÀsu. Zejmûˋna v tropech mohou ptûÀci migrovat jen velmi krûÀtkou vzdûÀlenost, jelikoéƒ zmány v habitatu a roánûÙm û¤hrnu srûÀéƒek se mohou zûÀsadná liéÀit v blûÙzkû§ch blûÙzkû§ch areûÀlech a ptûÀké₤m tak staáûÙ migrovat jen do sousednûÙ oblasti.[62] PéûÙkladem migranta na krûÀtkou vzdûÀlenost je strnadec zpávnû§, kterû§ se zdréƒuje mezi hnûÙzdiéÀti na hranicûÙch Spojenû§ch stûÀté₤ a Kanady, a zimoviéÀti ve stéednûÙch a jiéƒnûÙch áûÀstech Spojenû§ch stûÀté₤.[64] PéûÙkladem evropskûˋho migranta na krûÀtkou vzdûÀlenost mé₤éƒe bû§t kavka obecnûÀ, kterûÀ odlûˋtûÀ na zimu do jiéƒnûÙ Francie a karpatskûˋ kotliny.[65] Migranti na krûÀtkou vzdûÀlenost hnûÙzdûÙcûÙ v áesku tûÀhnou hlavná do zûÀpadnûÙ Evropy, zatûÙmco do áeska tûÀhnou ptûÀci ze severnûÙ Evropy váetná Polska.[66] Obecná platûÙ, éƒe migranti na krûÀtkou vzdûÀlenost tûÀhnou v menéÀûÙch hejnech neéƒ migranti na dlouhou vzdûÀlenost.[22]
Migrace na dlouhou vzdûÀlenost
Migrace na dlouhou vzdûÀlenost péedstavuje tah, báhem kterûˋho ptûÀci péekonûÀvajûÙ velkou geografickou péekûÀéƒku, jako je moée ái oceûÀn, migrujûÙ na jinû§ kontinent nebo péekraáujûÙcûÙ vûÙce podnebnû§ch pûÀsem.[63][61]
Tahovûˋ cesty satelitná sledovanû§ch béehouéÀé₤ rudû§ch. Mapa zobrazuje zemákouli s áarami péedstavujûÙcûÙ uletánou trasu béehouéÀé₤ z Novûˋho Zûˋlandu na Korejskû§ poloostrov z Novûˋho Zûˋlandu na sever. BéehouéÀi rudûÙ jsou rekordmani v nejdeléÀûÙm nepéetréƒitûˋm migraánûÙm letu (11 dnûÙ a 12 200 km)
JednûÙm z nejtypiátájéÀûÙch dûÀlkovû§ch migranté₤ je vlaéÀtovka obecnûÀ, jejûÙéƒ evropskûÀ populace (subsp. rustica) podnikûÀ kaéƒdoroánûÙ migraci z Evropy do africkû§ch, arabskû§ch a asijskû§ch tropé₤.[67] Dûˋlka trasy, kterou dûÀlkovûÙ migranti kaéƒdoroáná urazûÙ, mé₤éƒe jûÙt do tisûÙcé₤ i desetitisûÙcé₤ kilometré₤. JespûÀci rezavûÙ kaéƒdû§ rok lûˋtajûÙ z hnûÙzdiéÀéË v arktickûˋ Kanadá do zimoviéÀéË na jiéƒnûÙm cûÙpu JiéƒnûÙ Ameriky, péiáeméƒ urazûÙ aéƒ 6500 km báhem jedinûˋho nepéetréƒitûˋho letu.[68] Rekordmanem v nejdeléÀûÙm potvrzenûˋm nepéetréƒitûˋm migraánûÙm letu je véÀak béehouéÀ rudû§ (subsp. baueri), u kterûˋho byla namáéena migraánûÙ cesta z AljaéÀky na Novû§ Zûˋland dlouhûÀ 12 200 km. Let trval 11 dnûÙ a jedinec letál rychlostûÙ aéƒ 89 km/h. Tento vû§kon béehouéÀovi vyslouéƒil zûÀpis v Guinnessová knize rekordé₤ coby ãnejdeléÀûÙ nepéetréƒitûÀ ptaáûÙ migraceã.[69]
Vé₤bec nejdeléÀûÙ vzdûÀlenost mezi hnûÙzdiéÀtám a zimoviéÀtám ze véÀech zvûÙéat kaéƒdoroáná péekonûÀvûÀ rybûÀk dlouhoocasû§. RybûÀci hnûÙzdûÙ v Arktidá a zimujûÙ v antarktickû§ch oblastech. NejednûÀ se véÀak o nepéetréƒitou migraci jako u béehouéÀe rudûˋho; kaéƒdû§ migraánûÙ let trvûÀ kolem 2ã3 másûÙcé₤ a rybûÀci báhem náj áasto zastavujûÙ na odpoáinkovû§ch stanoviéÀtûÙch, aby nabrali sûÙly. PtûÀci báhem péesunu mezi hnûÙzdiéÀtám a zimoviéÀtám nalûˋtajûÙ kolem 25 tisûÙc kilometré₤. Za celû§ rok nalûˋtajûÙ i pées 80 000 km.[70] Podobná se na svû§ch cestûÀch zastavujûÙ i daléÀûÙ migranti, péiáeméƒ obáas kombinujûÙ péûÙmûˋ lety bez zastûÀvky a pomalejéÀûÙ lety s mnoéƒstvûÙm odpoáinku. PéûÙkladem mé₤éƒe bû§t lejsek bálokrkû§, kterû§ pozvolna migruje ze stéednûÙ a vû§chodnûÙ Evropy aéƒ na okraj StéedozemnûÙho moée, odkud lejsci tûÀhnou i 2ã3 dny v kuse do Afriky.[71]
HnûÙzdiéÀtá (zelená) a zimoviéÀtá (modée) báloéité₤ éÀedû§ch jsou ojedinálûÀ v tom, éƒe i americkûÀ populace migruje do Afriky
OjedinálûÀ je migrace báloéité₤ éÀedû§ch ze SevernûÙ Ameriky. Báloéiti se do severoamerickû§ch konáin rozéÀûÙéili z Evropy nákdy v pleistocûˋnu. Stejná jako evropskûÀ populace, i ta severoamerickûÀ migruje do zimoviéÀéË v subsaharskûˋ Africe. JednûÀ se o jedinûˋho zpávnûˋho ptûÀka, kterû§ migruje ze severnûÙ áûÀsti zûÀpadnûÙ polokoule do Afriky. MigraánûÙ trasa jednûÙm smárem mûÀ kolem 14 500 km a báloéiti báhem nûÙ urazûÙ kolem 290 km denná. Zé₤stûÀvûÀ otûÀzkou, jak dokûÀéƒe tento 25 g vûÀéƒûÙcûÙ, samostatná migrujûÙcûÙ pávec û¤spáéÀná zdolat takto obrovskou vzdûÀlenost, a to zvlûÀéÀtá uéƒ v prvnûÙch másûÙcûÙch éƒivota.[72][73]
Jednámi z nejvátéÀûÙch migranté₤ jsou trubkonosûÙ. éada trubkonosû§ch hnûÙzdûÙ na jiéƒnûÙ polokouli a migruje na tu severnûÙ.[74] TrubkonosûÙ s oblibou pro svûˋ lety vyuéƒûÙvajûÙ dostupnûˋ vátry, takéƒe jejich migraánûÙ trasy áasto nejsou péûÙmoáarûˋ. Napé. u buééûÀké₤ temnû§ch byla namáéena migrace dlouhûÀ 64 tisûÙc kilometré₤, protoéƒe buééûÀci letáli krouéƒivû§mi a elipsovitû§mi pohyby z Novûˋho Zûˋlandu pées Japonsko a AljaéÀku do Kalifornie a potûˋ zpát na Novû§ Zûˋland.[75]
Vû§éÀkovûÀ migrace
ZvlûÀéÀtnûÙm typem migrace je vû§éÀkovûÀ migrace. JednûÀ se o péesun mezi hnûÙzdiéÀti ve vyéÀéÀûÙch polohûÀch do nûÙéƒe poloéƒenû§ch zimoviéÀéË, kde je pées zimu dostatek potravy.[76] Odhaduje se, éƒe vû§éÀkovou migraci vykonûÀvûÀ kolem 10 % ze véÀech druhé₤ ptûÀké₤.[77] Na Havaji aéƒ 30 % druhé₤ ptûÀké₤ migruje vû§éÀková a v kontinentûÀlnûÙ SevernûÙ Americe vûÙce neéƒ 20 % druhé₤.[78]
TetéûÙvek douglaskovû§ provozuje tzv. péevrûÀcenou vû§éÀkovou migraci: na zimu se stahuje do hor, zatûÙmco lûˋta trûÀvûÙ v niéƒéÀûÙch polohûÀch
Vû§éÀkovûÀ migrace u ptûÀké₤ nastûÀvûÀ hlavná v tropech, kde bû§vûÀ dostatek potravy po celû§ rok, a ptûÀci mohou nalûˋzt vhodnûˋ podmûÙnky pro zimu péesunem pouhû§ch nákolika set vû§éÀkovû§ch metré₤. Tropickûˋ oblasti majûÙ celoroánûÙ dostatek svátla, takéƒe ptûÀci nemajûÙ potéebu migrovat za deléÀûÙmi dny.[79][80] Typickû§m péûÙkladem vû§éÀkovû§ch migranté₤ jsou kolibéûÙci ã kolem jednûˋ átvrtiny druhé₤ kolibéûÙké₤ vykonûÀvûÀ vû§éÀkovou migraci.[76][81]
ZatûÙmco nákterûˋ druhy vykonûÀvajûÙ pouze vû§éÀkovou migraci (krûÀtkû§ vû§éÀkovû§ péesun v rûÀmci jednoho pohoéûÙ), jinûˋ druhy vykonûÀvajûÙ zûÀroveé i dûÀlkovou migraci. ExtrûˋmnûÙm péûÙkladem jsou pánice horskûˋ, kterûˋ hnûÙzdûÙ v HimûÀlaji a zimujûÙ v jiéƒnûÙ Indii.[82] KrajnûÙm péûÙkladem z opaánûˋho konce jsou tetéûÙvci douglaskovûÙ, kteéûÙ migrujûÙ v nákterû§ch péûÙpadech pouhû§ch 300 m. V Guinnessová knize rekordé₤ jsou zapsûÀni coby vykonavatelûˋ ãnejkratéÀûÙ ptaáûÙ migraceã. Tito tetéûÙvci, podobná jako nákterûˋ daléÀûÙ ptaáûÙ druhy, vykonûÀvajûÙ tzv. péevrûÀcenou vû§éÀkovou migraci, jelikoéƒ se na zimu péesouvajûÙ do vyéÀéÀûÙch poloh a na lûˋto sestupujûÙ do nûÙéƒe poloéƒenû§ch oblastûÙ.[83]
áûÀsteánûÀ migrace
V péûÙpadá taéƒnû§ch ptûÀké₤ nemusûÙ nutná migrovat véÀichni jedinci danûˋho druhu, ale jen áûÀst populace. V takovûˋm péûÙpadá se jednûÀ o tzv. áûÀsteánou migraci. áûÀsteánûÀ migrace je velmi áastûÀ; napé. v AustrûÀlii lze za áûÀsteáná taéƒnûˋ ptûÀky oznaáit aéƒ 44 % pávcé₤ a 32 % ostatnûÙch ptûÀké₤.[84] V TasmûÀnii aéƒ 80 % ze véÀech stáhovavû§ch ptûÀké₤ tvoéûÙ áûÀsteánûÙ migranti. U nákterû§ch tasmûÀnskû§ch druhé₤ bylo zjiéÀtáno, éƒe migrujûÙ pouze v nákterû§ch letech. V áûÀsti JiéƒnûÙ Ameriky, kterûÀ zasahuje do jiéƒnûÙ polokoule, je kolem 70 % stáhovavû§ch ptûÀké₤ pouze áûÀsteánû§mi migranty.[85] HlavnûÙm dé₤vodem áûÀsteánûˋho tahu je vnitrodruhovûÀ konkurence o potravnûÙ zdroje v zimoviéÀtûÙch, kde jsou potravnûÙ zdroje dostateánûˋ pouze pro áûÀst populace, takéƒe ta druhûÀ áûÀst se vydûÀvûÀ za potravou jinde. ZdûÀ se, éƒe áûÀsteánûÀ migrace nastûÀvûÀ hlavná v oblastech se stéedná mûÙrnû§mi zimami, tzn. kde v zimá nedochûÀzûÙ k téeskutû§m mrazé₤m, avéÀak ani k nijak vysokû§m teplotûÀm.[86]
ZvlûÀéÀtnûÙ typy tahu
RozdûÙl mezi tzv. migracûÙ typu ãleapfrogã a éetázovou migracûÙ
Leapfrog
éetázovûÀ migrace
ZvlûÀéÀtnûÙm typem migrace je tzv. ãleapfrogã (pojem z angliátiny, kde oznaáuje dátskou hru, báhem kterûˋ dûÙtá skûÀáe pées ohnutûÀ zûÀda jinûˋho dûÙtáte). Péi tomto typu migrace se uráitûˋ populace druhu, kterûˋ obû§vajûÙ vyéÀéÀûÙ zemápisnûˋ éÀûÙéky neéƒ ostatnûÙ populace, péesunou na zimu do oblastûÙ niéƒéÀûÙch zemápisnû§ch éÀûÙéek, péiáeméƒ s kaéƒdou migracûÙ péeletûÙ (ãpéeéÀkoáûÙã) pées ostatnûÙ populace, kterûˋ jsou buá stûÀlûˋ, nebo migrujûÙ pouze na krûÀtkou vzdûÀlenost.[87] Klasickû§m péûÙkladem migrace typu ãleapfrogã je strnadec kaéÀtanovû§ ze SevernûÙ Ameriky.[88]
Vedle ãleapfrogã se rozliéÀuje jeéÀtá tzv. éetázovûÀ migrace (angl. chain migration), péi kterûˋ jiéƒnûÙ i severnûÙ populace druhu tûÀhnou stejnû§m smárem a urazûÙ zhruba stejnou vzdûÀlenost, takéƒe nakonec skonáûÙ ve stejnûˋm poéadûÙ, coéƒ je zûÀsadnûÙ rozdûÙl oproti ãleapfrogã migraci. Tento typ tahu nastûÀvûÀ napé. u rorû§sé₤ obecnû§ch.[89]
NoánûÙ migrace
VátéÀina druhé₤ ptûÀké₤ migruje v noci.[90] OstatnûÙ ptûÀci migrujûÙ buá ve dne, nebo ve dne i v noci.[91] K noánûÙm migranté₤m patéûÙ i dennûÙ druhy, tzn. ptûÀci, kteéûÙ jsou jinak aktivnûÙ ve dne. Dravci a plachtûÙcûÙ druhy se noánûÙm leté₤m vyhû§bajûÙ, protoéƒe báhem chladnû§ch nocûÙ se netvoéûÙ dostatek termickû§ch vátré₤. U ostatnûÙch druhé₤ nenûÙ zcela zéejmûˋ, podle jakûˋho klûÙáe se rozhodli pro noánûÙ migraci. Svou roli patrná bude hrûÀt celûÀ éada faktoré₤ od fyziologickû§ch po ty environmentûÀlnûÙ.[92] K nespornû§m vû§hodûÀm noánûÙ migrace patéûÙ relativná stabilnûÙ vátry, moéƒnost navigace podle hvázd, vátéÀûÙ mnoéƒstvûÙ dennûÙho svátla na krmenûÙ, efektivnûÙ vyhnutûÙ se predûÀtoré₤m nebo niéƒéÀûÙ teploty a vyéÀéÀûÙ vlhkost, coéƒ sniéƒuje riziko dehydratace.[93][94]
Hejno éÀpaáké₤ s áûÙhajûÙcûÙm dravû§m ptûÀkem (vpravo)
Nákterûˋ druhy predûÀtoré₤ se dokûÀzaly adaptovat na noánûÙ migraci ptûÀké₤, ze kterûˋ táéƒûÙ. K támto predûÀtoré₤m patéûÙ ostéûÙéƒ jiéƒnûÙ, kterû§ hnûÙzdûÙ ve StéedozemûÙ. U tohoto dravûˋho ptûÀka dochûÀzûÙ k pozdnûÙmu zahnûÙzdánûÙ (áervenecãéûÙjen), coéƒ je adaptace na hojnost potravy v tomto obdobûÙ. Tu pro ostéûÙéƒe zajiéÀéËujûÙ noánûÙ ptaáûÙ migranti (hlavná menéÀûÙ pávci) mezi Evropou a Afrikou, kteéûÙ pées hnûÙzdiéÀtá ostéûÙéƒé₤ na podzim hojná prolûˋtajûÙ.[95][96] K daléÀûÙm péûÙkladé₤m éƒivoáicha, kterû§ se adaptoval na noánûÙ migrace, patéûÙ netopû§r obrovskû§. Obecná platûÙ, éƒe jakûˋkoliv druhy netopû§ré₤ û¤toáûÙ na ptûÀky jen zcela vû§jimeáná, avéÀak éÀpanálskûˋ populace netopû§ré₤ obecnû§ch pravidelná konzumujûÙ menéÀûÙ migrujûÙcûÙ ptûÀky, kteéûÙ tudy na jaée a na podzim prolûˋtajûÙ. Netopû§éi obrovéÀtûÙ lovûÙ zejmûˋna menéÀûÙ druhy migrujûÙcûÙch pávcé₤ (napé. budnûÙáky lesnûÙ, áervenky obecnûˋ), kterûˋ patrná napadajûÙ péûÙmo za letu.[97]
ZatûÙmco dennûÙ migranti zaáûÙnajûÙ tah zrûÀna, noánûÙ migranti vylûˋtûÀvajûÙ naveáer. Pokud je to moéƒnûˋ, dosedajûÙ koncem noci, nicmûˋná napé. péi péeletu velkû§ch vodnûÙch ploch jsou druhy neuzpé₤sobenûˋ pobytu na vodá péinuceny pokraáovat v letu i za dne.[98] HlavnûÙ nevû§hodu noánûÙho tahu péedstavuje masivnûÙ redukce spûÀnku, jelikoéƒ ptûÀci v noci tûÀhnou a ve dne shûÀnájûÙ potravu. Ke krûÀtkûˋmu héadovûÀnûÙ mé₤éƒe dochûÀzet v pozdnûÙm odpoledni tásná péed poáûÀtkem tahu,[99] avéÀak nákteéûÙ ptûÀci nespûÙ vé₤bec (podrobnáji viz kapitola SpûÀnek). KrûÀtkûˋ odpolednûÙ ãschrupnutûÙã nastûÀvûÀ v dobá migrace i u noánûÙch migranté₤ chovanû§ch v zajetûÙ.[90]
NoánûÙ migranti báhem letu vydûÀvajûÙ áastûÀ krûÀtkûÀ kontaktnûÙ volûÀnûÙ. Tato volûÀnûÙ slouéƒûÙ k orientaci ptûÀké₤ v rûÀmci hejna, udréƒovûÀnûÙ vhodnû§ch rozestupé₤ i prevenci srûÀéƒek s ostatnûÙmi ptûÀky. ZdûÀ se, éƒe kontaktnûÙ volûÀnûÙ jsou dé₤leéƒitûÀ hlavná pro juvenilnûÙ jedince, kteéûÙ se báhem noánûÙch tahé₤ vokûÀlná projevujûÙ áastáji neéƒ dospálci.[100] Péi nejmenéÀûÙm u nákterû§ch druhé₤ majûÙ noánûÙ zvuky zakû°dovanou informaci o pohlavûÙ jedince a jeho identitá.[101]
Evoluce
U velkû§ch, sociûÀlná zaloéƒenû§ch druhé₤ jako jsou husy hraje sociûÀlnûÙ uáenûÙ tahovûˋ cesty vátéÀûÙ roli, neéƒ u menéÀûÙch druhé₤ ptûÀké₤ (na obrûÀzku hejno migrujûÙcûÙch berneéÀek bálolûÙcûÙch)
Migrace se u jednotlivû§ch ptaáûÙch taxoné₤ vyvinula dynamicky a nezûÀvisle na sobá.[102] MigraánûÙ chovûÀnûÙ je v ptûÀcûÙch geneticky zafixovûÀno a sociûÀlnûÙ uáenûÙ hraje nákdy sice dé₤leéƒitou, avéÀak druhotnou roli. JednûÙm z hlavnûÙch (negenetickû§ch) argumenté₤ pro genetickou podporu migrace je to, éƒe áerstvá narozenûÙ ptûÀci odlûˋtajûÙ z hnûÙzdiéÀéË do svû§ch zimoviéÀéË jeéÀtá péed zhoréÀenûÙm éƒivotnûÙch podmûÙnek v hnûÙzdiéÀtûÙch a bez podpory gené₤ by postrûÀdali motivaci pro tak energeticky nûÀroánou a riskantnûÙ aktivitu jako je migraánûÙ let. Negenetickûˋ faktory péesto mohou hrûÀt uráitou roli, jejûÙéƒ dé₤leéƒitost zûÀvisûÙ na druhu. NapéûÙklad nákterûˋ dlouho éƒijûÙcûÙ sociûÀlnûÙ druhy jako husy, áûÀpi, jeéûÀbi nebo dravci migrujûÙ v menéÀûÙch skupinûÀch vedenû§ch nejzkuéÀenájéÀûÙmi jedinci, kteéûÙ trasu jiéƒ nákolikrûÀt absolvovali. Naproti tomu menéÀûÙ taéƒnûÙ ptûÀci jako jsou pávci majûÙ tahovou cestu péevûÀéƒná vrozenou.[102][103][104]
Pod silnû§mi selekánûÙmi tlaky mé₤éƒe dojûÙt ke zmáná migraánûÙch genetickû§ch znaké₤ jiéƒ báhem nákolika generacûÙ. Péi pokusech s umále odchovanû§mi pánicemi áernohlavû§mi bylo zjiéÀtáno, éƒe tito pávci péi selektivnûÙm kéûÙéƒenûÙ v zajetûÙ opozdili odlet svûˋ podzimnûÙ migrace o vûÙce neéƒ tû§den v pré₤báhu pouhû§ch dvou generacûÙ.[105] Stejnûˋmu druhu se geneticky zafixovala i zmána tahovûˋ cesty jiéƒ báhem 30 let.[106]
K vû§voji migrace doéÀlo postupná ã ptûÀci nejdéûÙve hledali vhodnájéÀûÙ podmûÙnky pro éƒivot, naáeéƒ se postupná zaáali vydûÀvat dûÀle a dûÀle.[107][108] Tato skuteánost vzbuzuje otûÀzku, zda se ptûÀci vydûÀvali hledat vhodnájéÀûÙ zimoviéÀtá, áûÙméƒ by si zlepéÀili vyhlûÙdky na péeéƒitûÙ báhem zimnûÙho obdobûÙ, nebo se vydûÀvali hledat vhodnájéÀûÙ podmûÙnky pro zahnûÙzdánûÙ, coéƒ by zvyéÀovalo éÀance jejich reprodukánûÙ û¤spáéÀnosti. NejnovájéÀûÙ ornitologickûˋ vû§zkumy z poáûÀtké₤ 21. stoletûÙ se péiklûÀnûÙ k tomu, éƒe ptaáûÙ migrace se vyvinula postupnû§m objevovûÀnûÙm novû§ch zimoviéÀéË.[109] Dlouhodobá je totiéƒ pro ptûÀky vû§hodnájéÀûÙ várnost hnûÙzdiéÀti neéƒ zimoviéÀti.[109][110][111] Pokud ptûÀci hnûÙzdûÙ v oblasti s dostatkem zdrojé₤ po celû§ rok, nedojde k tvorbá selekánûÙch tlaké₤, nûÀsledkem kterû§ch se ptûÀci zaánou poohlûÙéƒet po novû§ch oblastech s péûÙhodnájéÀûÙmi éƒivotnûÙmi, resp. potravnûÙmi podmûÙnkami.[110]
Jedna z moéƒnû§ch péûÙáin vû§voje migraánûÙch nûÀvyké₤ ptûÀké₤ péedstavujûÙ glaciûÀlnûÙ cykly, kterû§ch se na Zemi vystéûÙdalo jen za poslednûÙch 2,5 milioné₤ let kolem 20. S rozéÀiéovûÀnûÙm ledovcé₤ báhem kaéƒdûˋ doby ledovûˋ smárem od polûÀrnûÙch oblastûÙ do niéƒéÀûÙch zemápisnû§ch éÀûÙéek dochûÀzelo k postupnûˋmu vytlaáovûÀnûÙ ptûÀké₤ a jejich hnûÙzdiéÀéË na jih; naopak s û¤stupem ledovcé₤ dochûÀzelo k odhalovûÀnûÙ novû§ch û¤rodnû§ch oblastûÙ, kterûˋ se ptûÀké₤m zpéûÙstupnily.[111] S roztahovûÀnûÙm ledovcé₤ nedoéÀlo k pouhûˋmu zkrûÀcenûÙ dûˋlky migraánûÙch tras ã u nákterû§ch druhé₤ doéÀlo k û¤plnûˋmu vynechûÀnûÙ migrace.[112]
Fyziologie a kontrola
Migrace je spojena s rychlû§m éƒivotnûÙm tempem ã migraánûÙ druhy oproti stûÀlû§m druhé₤m vykazujûÙ vyéÀéÀûÙ plodnost, dospûÙvajûÙ déûÙve a neéƒijûÙ tak dlouho. U áûÀsteáná taéƒnû§ch ptûÀké₤ je tento rozdûÙl o náco menéÀûÙ.[8]
NaáasovûÀnûÙ
Pánice slavûÙkovûÀ vykazujûÙcûÙ tahovû§ neklid v laboratornûÙch podmûÙnkûÀch
é§ivot ptûÀké₤ se éûÙdûÙ vrozenû§m endogennûÙm celoroánûÙm (cirkanuûÀlnûÙm) rytmem. Tento geneticky fixovanû§ rytmus seéûÙzenû§ podle dûˋlky svátla báhem dne uráuje péibliéƒnû§ áas jarnûÙ i podzimnûÙ migrace.[93][113][114] DûÀle majûÙ na naáasovûÀnûÙ migrace vliv i genetickûÀ vû§bava, poáasûÙ, sociûÀlnûÙ stimuly, biologickûÀ zdatnost, teplota nebo dostupnost potravy.[93]
Tásná péed migracûÙ ptûÀci zaáûÙnajûÙ vykazovat fyziologickû§ proces zvanû§ tahovû§ neklid, v odbornûˋ literatuée oznaáovûÀn námeckû§m vû§razem Zugunruhe. Tahovû§ neklid se vyznaáuje zvû§éÀenou aktivitou a nervozitou nákdy aéƒ hraniáûÙcûÙ s û¤zkostûÙ. U ptûÀké₤ v zajetûÙ se tento fenomûˋn projevuje vysokou noánûÙ aktivitou, nespavostûÙ, neustûÀlû§m poskakovûÀnûÙm a téepotûÀnûÙm kéûÙdel. Dûˋlka, po kterou ptûÀci v zajetûÙ vykazujûÙ tahovû§ neklid, je péibliéƒná stejnûÀ jako dûˋlka migrace druhu ve volnûˋ péûÙrodá.[99] V dobá tahovûˋho neklidu se ptûÀci v zajetûÙ orientujûÙ smárem k destinaci migraánûÙ trasy populace ve volnûˋ péûÙrodá.[113]
Orientace a navigace
PtûÀci pro navigaci vyuéƒûÙvajûÙ magnetickûˋho pole Zemá, olfaktorickûˋ (áichovûˋ) vnûÙmûÀnûÙ, vizuûÀlnûÙ prvky v terûˋnu i polohu Slunce a hvázd.[115] VypadûÀ to, éƒe ptûÀci báhem navigace pouéƒûÙvajûÙ kombinaci véÀech táchto prvké₤, kterûˋ mezi sebou porovnûÀvajûÙ a vyhodnocujûÙ. ZatûÙmco pro nákterûˋ druhy mé₤éƒe bû§t hlavnûÙm navigaánûÙm prvkem magnetickûˋ pole Zemá, u jinû§ch druhé₤ to je áichovûÀ kognitivnûÙ mapa (napé. trubkonosûÙ).[116][117]
I pées intenzivnûÙ vádeckû§ vû§zkum a éadu praktickû§ch pokusé₤ zé₤stûÀvûÀ péesnû§ zpé₤sob navigace a orientace ptûÀké₤ z velkûˋ áûÀsti nevysvátlen.[117] Zejmûˋna noánûÙ migrace dennûÙch druhé₤ vzbuzuje otûÀzku, jakû§m zpé₤sobem se ptûÀci v noci navigujûÙ, protoéƒe jejich noánûÙ vize áasto nemusûÙ bû§t o nic lepéÀûÙ neéƒ noánûÙ vize álováka.[118] Podle laboratornûÙch experimenté₤ se noánûÙ migranti mohou navigovat pomocûÙ magnetickûˋho pole Zemá, kterûˋ patrná majûÙ schopnost vidát skrze zvlûÀéÀtnûÙ fotoreceptory zvanûˋ kryptochromy. Tyto lûÀtky se vyskytujûÙ v sûÙtnici nákterû§ch ptûÀké₤, a patrná reagujûÙ na elektromagnetickûˋ vlnánûÙ zemskûˋho magnetickûˋho pole.[119][120]
MasivnûÙ houfovûÀnûÙ áûÀpé₤ bûÙlû§ch nad Istanbulem
Zatoulanci
Obáas se stane, éƒe se stáhovavûÙ ptûÀci zatoulajûÙ do konáin, kde se báéƒná nevyskytujûÙ. V takovûˋm péûÙpadá jsou oznaáovûÀni za zatoulance.[53] V krajnûÙch péûÙpadech se mohou ocitnout i tisûÙce kilometré₤ od báéƒnûˋho mûÙsta vû§skytu.[121] NejáastájéÀûÙmi dé₤vody táchto zûÀleté₤ bû§vûÀ nepéûÙznivûˋ poáasûÙ (napé. silnû§ vûÙtr nebo snaha vyhnout se bouéi) nebo dezorientace ptûÀké₤.[122] Péi nejmenéÀûÙm nákteéûÙ ptûÀci se dokûÀéƒou s jevy poáasûÙ vyrovnat. U orlovcé₤ éûÙánûÙch a váelojedé₤ lesnûÙch bylo zjiéÀtáno, éƒe staréÀûÙ jedinci báhem letu umûÙ vykompenzovat silnûˋ vátry zmánánûÙm kurzu, takéƒe vûÙtr neovlivnûÙ smár jejich letu tak jako mladéÀûÙ jedince.[123] NákteéûÙ ptûÀci dokûÀéƒou silnû§ch vátré₤ naopak vyuéƒûÙt ve své₤j prospách. MenéÀûÙ druhy ptûÀké₤ ze severnûÙ Ameriky vyuéƒûÙvajûÙ severozûÀpadnûÙ vátry k podpoée leté₤ pées zûÀpadnûÙ Atlantik smárem do JiéƒnûÙ Ameriky.[124]
ZvlûÀéÀtnûÙ péûÙpad zatoulûÀnûÙ ptûÀké₤ péedstavuje tzv. reverznûÙ migrace (reverse migration). Tento fenomûˋn oznaáuje situaci, kdy ptûÀci migrujûÙ opaánû§m smárem, neéƒ kterû§m by normûÀlná máli letát.[121] ReverznûÙ migrace bû§vajûÙ spojovûÀny mj. s extrûˋmnûÙmi vû§kyvy poáasûÙ nebo geografickû§mi bariûˋrami, na kterûˋ narazûÙ juvenilnûÙ, prvná migrujûÙcûÙ jedinci.[125] BritéÀtûÙ ornitologovûˋ James Gilroy a Alexander Charles Lees nicmûˋná upozornili na to, éƒe u ptûÀku sice dochûÀzûÙ k dezorientaci nejáastáji opaánû§m smárem, nicmûˋná ptûÀci mohou zamûÙéit i jinû§mi smáry. Tyto nûÀhodnûˋ rozptyly ptûÀké₤ jsou podle nich zpé₤sobeny genetickû§mi abnormalitami.[126]
DaléÀûÙm fenomûˋnem migraánûÙch zatoulancé₤ je tzv. péestéelenûÙ (overshooting), ke kterûˋmu dochûÀzûÙ, pokud ptûÀci péi tahu z jakûˋhokoliv dé₤vodu nepéistanou v mûÙstá zimoviéÀtá ái hnûÙzdiéÀtá a pokraáujûÙ v letu dûÀle. Dé₤vody mohou bû§t ré₤znûˋ, mj. se mé₤éƒe jednat o péehlûˋdnutûÙ ostatnûÙch jedincé₤ tûˋhoéƒ druhu nebo nerozpoznûÀnûÙ vhodnûˋho biotopu, takéƒe migrujûÙcûÙ ptûÀci letûÙ dûÀle a dûÀle.[126]
ZatoulanûÙ jedinci nemusûÙ nutná znamenat pro ptûÀky problûˋm ã jsou zdokumentovûÀny péûÙpady, kdy ãzatoulanostã ptûÀké₤ vedla k objevenûÙ novû§ch tahovû§ch cest, zimoviéÀéË nebo hnûÙzdiéÀéË.[127][128] ZatoulanûÙ ptûÀci tak mohou hrûÀt vû§znamnou roli v areûÀlu rozéÀûÙéenûÙ ptûÀké₤.[122]
AdaptaánûÙ mechanismy
BéehouéÀi patéûÙ k ptûÀké₤m s nejdeléÀûÙ migraánûÙ cestou v jednom tahu i k ptûÀké₤m s nejvátéÀûÙm pomárovû§m nûÀré₤stem tuku vzhledem k objemu tála v dobá péed migraánûÙm letem (na obrûÀzku béehouéÀ rudû§)
U stáhovavû§ch ptûÀké₤ se vyvinula éada fyziologickû§ch, hormonûÀlnûÙch i neurologickû§ch adaptacûÙ, kterûˋ jim pomûÀhajûÙ vyrovnat se s extrûˋmnûÙ povahou migrace. K támto adaptacûÙm patéûÙ hlavná schopnost tála navû§éÀit zûÀsoby tuku péed migracûÙ, kontrola spûÀnku, transformace vniténûÙch orgûÀné₤ nebo péepeéovûÀnûÙ.
ZûÀsobovûÀnûÙ tukem
MigraánûÙ druhy jsou uzpé₤sobeny jak tvorbá tukovû§ch zûÀsob péed odletem, tak obdobûÙm pé₤stu (napé. báhem dlouhû§ch migraánûÙch leté₤). Tásná péed poáûÀtkem tahu mohou tukovûˋ zûÀsoby naré₤st aéƒ na 50ã60 % celkovûˋho tálesnûˋho objemu,[129] avéÀak vû§éÀe tukovû§ch zûÀsob zûÀvisûÙ na dûˋlce migraánûÙ trasy i na tom, jak áasto nebo jestli vé₤bec se ptûÀci chystajûÙ podniknout tahovûˋ zastûÀvky a doplnit zûÀsoby. V dobá krmenûÙ péed migraánûÙm letem dochûÀzûÙ u metabolismu ptûÀké₤ k hormonûÀlnûÙm zmánûÀm, kterûˋ jim umoéƒnûÙ efektivnáji naklûÀdat s pozéenû§m jûÙdlem a uklûÀdat ho ve formá tuké₤.[130] Hormony (resp. hormon ghrelin, kterû§ je spojovûÀn s apetitem) patrná rozhodujûÙ za ptûÀky i to, zda jsou jejich zûÀsoby tuku jiéƒ dostateánûˋ pro nadchûÀzejûÙcûÙ let.[131]
ZûÀsoby tuku jsou uloéƒeny v tukovû§ch tkûÀnûÙch ve formá energeticky bohatû§ch triglyceridé₤. OxidacûÙ táchto mastnû§ch kyselin báhem letu dochûÀzûÙ ke tvorbá a distribuci potéebnûˋ energie do svalé₤ a cûˋvnûÙ soustavy.[132]
Transformace vniténûÙch orgûÀné₤
U taéƒnû§ch ptûÀké₤ dochûÀzûÙ v pré₤báhu roku ke zmánûÀm velikosti vniténûÙch orgûÀné₤. Péed migraánûÙm letem nastûÀvûÀ smréÀtánûÙ tách vniténûÙch orgûÀné₤, kterûˋ báhem energeticky vyáerpûÀvajûÙcûÙho letu jednoduéÀe ãnejsou potéebaã. K tám patéûÙ hlavná orgûÀny zaéƒûÙvacûÙ soustavy, kterûˋ se mohou smrsknout o vûÙce neéƒ 50 % (ledviny, jûÀtra, stéeva).[133][134] Naopak u orgûÀné₤ klûÙáovû§ch k letu (prsnûÙ svaly, srdce, plûÙce) mé₤éƒe dojûÙt k jejich roztaéƒenûÙ. OrgûÀny obnovûÙ svou pé₤vodnûÙ velikost po péûÙletu do zimoviéÀtá nebo na odpoáinkovûˋ stanoviéÀtá.[135][136][137]
Obnova orgûÀné₤ na jejich pé₤vodnûÙ velikost trvûÀ i nákolik dnûÙ, péiáeméƒ hlavná péi tahovû§ch zastûÀvkûÀch ptûÀci mohou kompenzovat zmenéÀenûˋ orgûÀny trûÀvicûÙho û¤strojûÙ pomocûÙ prodlouéƒenûˋ doby zaéƒûÙvûÀnûÙ.[133]
Rorû§s velkû§ dokûÀéƒe ve vzduchu strûÀvit i pé₤l roku
SpûÀnek
SpûÀnek ptûÀké₤, zvlûÀéÀtá báhem dlouhû§ch migraánûÙch leté₤, zé₤stûÀvûÀ z velkûˋ áûÀsti nevysvátlen.[93] Je potvrzeno, éƒe celûÀ éada ptûÀké₤ dokûÀéƒe spûÀt za letu. U fregatek obecnû§ch bylo prokûÀzûÀno, éƒe dovedou spûÀt báhem letu s jednou (tzv. spanûÙ na pé₤l mozku) nebo obáma hemisfûˋrami. ZatûÙmco báhem pobytu na souéÀi fregatky prospûÙ kolem 12 hodin denná, báhem dlouhû§ch leté₤ spûÙ v pré₤máru pouze kolem 40 minut denná, aniéƒ by to bylo poznat na jejich tálesnûˋm vû§konu ái pozornosti. NenûÙ û¤plná zéejmûˋ, jak fregatky dokûÀéƒou vystaáit po dobu nákolika tû§dné₤ i másûÙcé₤ s tak malû§m mnoéƒstvûÙm spûÀnku; vysvátlenûÙ bude patrná souviset s neurochemickû§mi procesy v mozku.[138]
U strnadcé₤ bálopûÀsû§ch bylo pokusy vyzkoumûÀno, éƒe strnadci dokûÀéƒou báhem taéƒnûˋho obdobûÙ zredukovat mnoéƒstvûÙ svûˋho spûÀnku o dvá téetiny, aniéƒ by se snûÙéƒila péesnost jejich reakcûÙ na péedklûÀdanûˋ û¤koly. Naproti tomu v dobá mimo taéƒnûˋ obdobûÙ byla reakce strnadcé₤ na péedklûÀdanûˋ û¤koly rapidná snûÙéƒena jiéƒ po jednom dni s redukovanû§m mnoéƒstvûÙm spûÀnku.[99]Rorû§si velcûÙ dokûÀéƒûÙ spûÀt za letu a zé₤stat tak ve vzduchu po dobu nejmûˋná pé₤l roku.[139]
PéepeéovûÀnûÙ
U taéƒnû§ch druhé₤ lze rozliéÀit dvá nejáastájéÀûÙ péepeéovacûÙ strategie: k péepeéovûÀnûÙ dochûÀzûÙ buá po obdobûÙ rozmnoéƒovûÀnûÙ v oblasti hnûÙzdiéÀéË péed migraánûÙm letem, nebo (áastáji) aéƒ po péûÙletu do zimoviéÀéË. Obá strategie péitom majûÙ svûˋ vû§hody: áerstvá péepeéenûÙ jedinci péed migraánûÙm letem majûÙ novûˋ peéûÙ, kterûˋ pomûÀhûÀ zefektivnit let; naopak post-migraánûÙ péepeéenûÙ pomé₤éƒe s obnovou peéûÙ, kterûˋ se péi tahu mé₤éƒe znaáná obnosit. Nadto existuje celûÀ éada daléÀûÙch ré₤znû§ch variacûÙ péepeéovacûÙ strategie. NákteéûÙ ptûÀci péepeéujûÙ hned dvakrûÀt roáná (napé. budnûÙáek vátéÀûÙ[140]), éada druhé₤ péepeéuje báhem migrace (napé. rûÀkosnûÙk velkû§, leséûÀáek lejsáûÙ), péiáeméƒ nákterûˋ z táchto druhé₤ péepeéujûÙ jen nákterûˋ letky (napé. cvráilka éûÙánûÙ[141]).[142]
Migrace ptûÀké₤ v áesku a Evropá
Tabulka s vybranû§mi áeskû§mi druhy stáhovavû§ch ptûÀké₤ zobrazujûÙcûÙ dobu jejich péûÙletu a odletu do áeska
SchematickûÀ mapka zobrazujûÙcûÙ téi hlavnûÙ tahovûˋ cesty áeskû§ch stáhovavû§ch ptûÀké₤ do Afriky
PtûÀci nejáastáji odlûˋtajûÙ do oblasti StéedomoéûÙ, Pyrenejskûˋho a Apeninskûˋho poloostrova, nákterûˋ druhy véÀak odlûˋtajûÙ dûÀle do Afriky.[146] PtûÀci hnûÙzdûÙcûÙ v áesku, potaéƒmo stéednûÙ Evropá, odlûˋtajûÙ do svû§ch zimoviéÀéË téemi hlavnûÙmi tahovû§mi cestami:
jihozûÀpadnûÙ cestou pées Pyrenejskû§ poloostrov a Gibraltarskû§ pré₤liv;
jihovû§chodnûÙ cestou pées BalkûÀnskû§ poloostrov a Sû§rii;
apeninskou cestou vedoucûÙ pées Apeninskû§ poloostrov a SicûÙlii a ãnapéûÙmoã pées StéedozemnûÙ moée.[147]
ZatûÙmco prvnûÙ dvá cesty vedou podûˋl pevniny a snaéƒûÙ se tak vyhnout otevéenû§m plochûÀm, ta téetûÙ péekonûÀvûÀ vû§znamnou geografickou bariûˋru ã StéedozemnûÙ moée. DûÙky relativnûÙ ustûÀlenosti tahovû§ch cest i pravidelnosti migrace lze péedvûÙdat, kudy a kdy budou stáhovavûˋ druhy áeskem protahovat. Toho vyuéƒûÙvajûÙ odchytovûˋ stanice, kterûˋ krouéƒkujûÙ ptûÀky za û¤áelem vádeckûˋho vû§zkumu a poznûÀnûÙ migrace. NejaktivnájéÀûÙm krouéƒkovacûÙm stanoviéÀtám v áesku je odchytovûÀ stanice na áervenohorskûˋm sedle, kterûˋ tvoéûÙ péirozenû§ trichtû§é Jesenickûˋho masivu. Mezi lety 2010ã2018 bylo na áervenohorskûˋm sedle odchyceno a okrouéƒkovûÀno kolem 100 jedincé₤ 125 druhé₤ ptûÀké₤.[148]
V Evropskûˋ unii váetná áeska jsou stáhovavûˋ druhy chrûÀnány mj. Bonnskou û¤mluvou,[149] nicmûˋná i tak dochûÀzûÙ v nákterû§ch álenskû§ch stûÀtech k odstéelé₤m stáhovavû§ch druhé₤ v masovû§ch máéûÙtkûÀch pro ãsportã. Na Maltá, kudy kaéƒdoroáná protahuje kolem 40 milioné₤ ptûÀké₤, je kaéƒdoroáná pytlûÀky zastéeleno kolem 2 milioné₤ ptûÀké₤.[150] Podobná i na Kypru bylo jen v roce 2016 uloveno cca 2,3 milionu ptûÀké₤ (péedevéÀûÙm pávcé₤), kteéûÙ tvoéûÙ hlavnûÙ péûÙsadu do mûÙstnûÙ ãpochoutkyã zvanûˋ ambelopoulia.[151] Odhaduje se, éƒe ve StéedomoéûÙ kaéƒdoroáná hyne aéƒ 25 milioné₤ ptûÀké₤ nûÀsledkem pytlaáenûÙ.[152]
PtaáûÙ migrace a álovák
Historickûˋ perspektivy
Tichomoéskûˋ plavby
DvojitûÀ kûÀnoe z Fidéƒi; na podobnû§ch kûÀnoûÙch se pohybovaly tichomoéskûˋ nûÀrody napéûÙá Tichû§m oceûÀnem. Indicie ke smáru plaveb zûÙskûÀvali pozorovûÀnûÙm okolnûÙho prostéedûÙ, mimo jinûˋ patrná i migrace ptûÀké₤
ZmûÙnky o migraci ptûÀké₤ se objevujûÙ v orûÀlnûÙch i pûÙsemnû§ch tradicûÙch lidstva jiéƒ od pradûÀvna. PtaáûÙ migrace patrná hrûÀly dé₤leéƒitou roli v navigaci plaveb Polynûˋsané₤ a Mikronûˋsané₤, jiméƒ jiéƒ péed nákolika tisûÙci lety poskytovaly indicie o existenci pevniny, coéƒ podporujûÙ mnohûˋ legendy dochovanûˋ v orûÀlnûÙch tradicûÙch tichomoéskû§ch nûÀrodé₤. Napé. na ostrová Bougainville se traduje, éƒe péedci tamnûÙch obyvatel vydedukovali péûÙtomnost dosud neznûÀmûˋho ostrova pozorovûÀnûÙm tahu ptûÀké₤. Domorodci si vyhlûˋdli hvázdu, kterûÀ byla ve smáru jejich letu, nasedli do kûÀnoûÙ a vypluli ve smáru danûˋ hvázdy, dokud nenarazili na ostrov, kterû§ potûˋ osûÙdlili.[153] Podobnûˋ zmûÙnky se vyskytujûÙ i v daléÀûÙch legendûÀch. V samojskûˋ kultuée se traduje, éƒe bé₤h Tagaloa seslal svou dceru Sinu na Zemi ve formá ptûÀka Tuli, aby naéÀel pevninu.[154] Jako tuli se v samojéÀtiná oznaáuje éada druhé₤ brodivû§ch ptûÀké₤, mj. kulûÙk pacifickû§,[155] kterû§ migruje mezi arktickû§mi oblastmi a Novû§m Zûˋlandem.[156]
Vedle ostrové₤ a atolé₤ v TichomoéûÙ migrace ptûÀké₤ patrná pomohly Polynûˋsané₤m objevit i novozûˋlandskûˋ souostrovûÙ, kterûˋ péedstavuje jednu z poslednûÙch velkû§ch pevnin osûÙdlenû§ch lidmi. Na Novû§ Zûˋland kaéƒdoroáná migruje pées 20 milioné₤ ptûÀké₤, coéƒ by patrná jen táéƒko uniklo obyvatelé₤m polynûˋskû§ch ostrové₤, kteéûÙ jsou znûÀmûÙ vû§bornû§mi pozorovacûÙmi schopnostmi okolnûÙ péûÙrody.[154] JednûÙm z nejáastáji zmiéovanû§ch ptûÀké₤, kterû§ mohl slouéƒit jako ukazatel novozûˋlandskûˋ pevniny, je kukaáka dlouhoocasûÀ, kterûÀ kaéƒdoroáná migruje mezi hnûÙzdiéÀti na Novûˋm Zûˋlandu a zimoviéÀti v jiéƒnûÙm TichomoéûÙ.[154][157] Podobná i kukaáka nûÀdhernûÀ kaéƒdoroáná migruje mezi novozûˋlandskou pevninou a mikronûˋskû§mi ostrovy.[157]
ZûÀpadnûÙ tradice
I áûÀp na nebi znûÀ své₤j áas, hrdliáka, vlaéÀtovka a jeéûÀb dodréƒujûÙ áas svûˋho péûÙletu, ale mé₤j lid neznûÀ Hospodinovy éûÀdy.
K nejstaréÀûÙm pûÙsemnû§m zmûÙnkûÀm o migraci ptûÀké₤ v tradici ZûÀpadu patéûÙ HomûˋrovaIlliada z 8. stoletûÙ pé. n. l., kde je trojskûÀ armûÀda péirovnûÀvûÀna k tûÀhnoucûÙmu hejnu jeéûÀbé₤.[159] ZmûÙnky o migraci ptûÀké₤ lze nalûˋzt i ve Starûˋm zûÀkoná, konkrûˋtná v Knize Jû°b[160] nebo Knize JeremijûÀéÀ.[158] PtaáûÙ migracûÙ se zabû§vali staroéeátûÙ klasiátûÙ myslitelûˋ jako byli Hûˋrodotos, Hûˋsiodos a Aristotelûˋs.[161] Aristotelûˋs si mj. véÀiml, éƒe ptûÀci péed odletem ztloustnou. Tento filosof uvaéƒoval o téech moéƒnû§ch vysvátlenûÙch sezû°nnûÙho odletu ptûÀké₤: ptûÀci buá migrujûÙ do vzdûÀlenû§ch konáin, nebo se transformujûÙ do jinûˋ podoby (tzv. teorie trasmutace), nebo hibernujûÙ. Aristotelûˋs zavrhl migraci jako bizarnûÙ, avéÀak transmutacûÙ a hibernacûÙ se podrobnáji zabû§val.[162] Aristotelovo uvaéƒovûÀnûÙ o transmutaci bylo motivovûÀno tûÙm, éƒe zatûÙmco jeden druh odlûˋtûÀ na sever, jinû§ nedlouho potûˋ péilûˋtûÀ z jihu. Aristotelûˋs proto uvaéƒoval, éƒe se jednûÀ o tytûˋéƒ ptûÀky, kteéûÙ se véÀak péemánili na jinû§ druh. Tento éeckû§ velikûÀn propagoval i teorii hibernace, kterou pozdáji éÀûÙéili jeho nûÀsledovnûÙci. Podle hibernaánûÙ teorie se taéƒnûÙ ptûÀci na chladnûˋ obdobûÙ uchylujûÙ do jeskynûÙ, dutin stromé₤ ái dokonce do bahna moáûÀlé₤ a jezer, kde v zimnûÙm obdobûÙ hibernujûÙ.[163][164]
Migrace vs hibernace vlaéÀtovek
RybûÀéi vytahujûÙcûÙ ryby a hibernujûÙcûÙ vlaéÀtovky (Olaus Magnus, 1555)
Aristotelova teorie hibernace ptûÀké₤ se stala péevaéƒujûÙcûÙ teoriûÙ vysvátlenûÙ kaéƒdoroánûÙho tahu vlaéÀtovek na daléÀûÙ 2 tisûÙce let. V roce 1555 teorii ãposvátilã uppsalskû§ arcibiskupOlaus Magnus, kterû§ popsal, jak v severnûÙch moéûÙch rybûÀéi vytahujûÙ sûÙtá plnûˋ ryb a vlaéÀtovek.[165] Podle Magnuse staáilo vlaéÀtovky péemûÙstit do tepla vyhéûÀtûˋho domu, naáeéƒ vlaéÀtovky oéƒily, avéÀak nedlouho na to uhynuly.[161] Vedle hibernace jako moéƒnûˋho vysvátlenûÙ tahu ptûÀké₤ se uvaéƒovalo obáas i o jinû§ch teoriûÙch. SvûˋrûÀzná se s otûÀzkou vypoéûÀdal novoanglickû§ uáenec Charles Morton, kterû§ v roce 1694 sice sprûÀvná napsal, éƒe motivacûÙ pro migraci ptûÀké₤ jsou zmány v okolnûÙm prostéedûÙ a v dostupnost potravy, nicmûˋná navrhl, éƒe ptûÀci odlûˋtajûÙ na MásûÙc.[166]
PrvnûÙ nûÀznaky vyéeéÀenûÙ ãzûÀhady mizenûÙ vlaéÀtovekã se objevily na péelomu 18. a 19. stoletûÙ. Anglickû§ uáenec Thomas Bewick ve svûˋm zûÀsadnûÙm dûÙle A History of British Birds (Historie britskû§ch ptûÀké₤) z roku 1797 zmiéuje vû§pováá ãvelice inteligentnûÙho lodnûÙho kapitûÀnaã, kterû§ mezi ostrovy Mallorca a Menorca pozoroval velkûÀ hejna vlaéÀtovek mûÙéûÙcûÙch na sever. Bewick dûÀle zavrhl moéƒnost hibernace vlaéÀtovek ve vodá, coéƒ je podle náj chybnûˋ tvrzenûÙ zaloéƒenûˋ na skuteánosti, éƒe se vlaéÀtovky stahujûÙ na noc k vodnûÙm plochûÀm, kde héadujûÙ. Snad nejvátéÀûÙ péûÙnos Bewicka k vyéeéÀenûÙ otûÀzky mizenûÙ vlaéÀtovek bylo to, éƒe nákolik vlaéÀtovek odchytil a udréƒoval je v zajetûÙ v teple a suchu po nákolik britskû§ch zim, áûÙméƒ dokûÀzal, éƒe u nich nedochûÀzûÙ k hibernaci. Bewick uzavéel otûÀzku migrace tûÙm, éƒe vlaéÀtovky opouéÀtûÙ BritûÀnii jednoduéÀe potûˋ, co dojde k vyáerpûÀnûÙ jejich potravnûÙch zdrojé₤.[164][167]
Freska zobrazujûÙcûÙ vlaéÀtovky (neznûÀmû§ autor, Akrotiri, cca 1600-1500 pé. n. l.)
Moéƒnost hibernace vlaéÀtovek se dlouho jevila jako logickûÀ, protoéƒe byla znûÀma u nákterû§ch jinû§ch zvûÙéat jako jsou obojéƒivelnûÙci a plazi. Nákterûˋ kultury si vysvátlovaly mizenûÙ ptûÀké₤ po svûˋm. Dodnes se u nákterû§ch jihoevropskû§ch nûÀrodé₤ traduje, éƒe malûˋ ptûÀky péenûÀéÀûÙ pées StéedozemnûÙ moée vátéÀûÙ druhy.[168][169] Podobná se v tradicûÙch nákterû§ch kmené₤ pé₤vodnûÙch obyvatel americkûˋho kontinentu traduje, éƒe aljaéÀéÀtûÙ kolibéûÙci se péemiséËujûÙ na zimu na jih na hébetech berneéÀek velkû§ch, jiméƒ se zavrtajûÙ do peéûÙ.[168][170] PéesvádáenûÙ o hibernaci vlaéÀtovek véÀak bylo mezi péûÙrodovádci nadûÀle silná zakoéenáno. é vûˋdskû§ péûÙrodovádec Carl Linnûˋ v roce 1845 vypsal odmánu tomu, kdo vlaéÀtovky v bahná objevûÙ.[170] Americkû§ ornitolog Elliott Coues roce 1878 publikoval seznam hned 182 álûÀnké₤, kterûˋ se zabû§valy hibernacûÙ vlaéÀtovek. SûÀm Coues se k hibernaci stavál ponákud odmáéená, kdyéƒ napsal: ãNikdy jsem nic takovûˋho nevidál, ani neznûÀm nikoho, kdo to [hibernaci] vidál [...]ã.[162]
Pfeilstorch a prvnûÙ studia migrace
Pfeilstorch ze zoologickûˋ sbûÙrky Roztockûˋ univerzity
Vû§znamnû§m posunem v rozluéÀtánûÙ ãzûÀhadyã mizenûÙ ptûÀké₤ byl nûÀlez áûÀpa bûÙlûˋho s éÀûÙpem zabodnutû§m v krku. Tento jedinec byl odchycen v mástá Klû¥tz v dneéÀnûÙm severnûÙm Námecku v roce 1822 a poskytuje jeden z prvnûÙch solidnûÙch dé₤kazé₤ dûÀlkovûˋ migrace ptûÀké₤.[171] é ûÙp mál na dûˋlku kolem 80 cm a byl vyroben z africkûˋho déeva, áûÙméƒ bylo jasnûˋ, éƒe áûÀp byl zasaéƒen éÀûÙpem na û¤zemûÙ Afriky, odkud vykonal nákolika tisûÙcikilometrovû§ let aéƒ k Baltskûˋmu moéi, kde byl odchycen.[172] Tento áûÀp byl nazvûÀn pfeilstorch (z námeckûˋho pfeil, áili ãéÀûÙpã, a storch, ãáûÀpã). Jedinec byl vypreparovûÀn a stal se souáûÀstûÙ zoologickûˋ sbûÙrky Roztockûˋ univerzity.[171] K roku 2003 bylo evidovûÀno kolem 25 péûÙpadé₤ pfeilstorch.[172]
V pré₤báhu 19. stoletûÙ se nûÀzor vádcé₤ na kaéƒdoroánûÙ odlet ptûÀké₤ postupná zaáal péiklûÀnát na stranu migrace. Americkû§ ornitolog George Lowery kolem poloviny 20. stoletûÙ vyvinul techniku zaznamenûÀvûÀnûÙ noánûÙch migranté₤ skrze teleskopy namûÙéenûˋ k û¤pléku a poáûÙtûÀnûÙ proletánû§ch siluet.[173] KlûÙáovû§m krokem ke studiu migrace ptûÀké₤ byl vynûÀlez krouéƒkovûÀnûÙ, se kterû§m péiéÀel dûÀnskû§ ornitolog Hans Mortensen na péelomu 19. a 20. stoletûÙ. V prvnûÙch desetiletûÙch 20. stoletûÙ zaáaly vychûÀzet prvnûÙ studie publikujûÙcûÙ vû§sledky krouéƒkovacûÙch projekté₤.[174]
ModernûÙ metody studia migrace
Ke klûÙáovû§m modernûÙm vádeckû§m metodûÀm studia migrace stûÀle patéûÙ krouéƒkovûÀnûÙ ptûÀké₤. JednûÀ se o levnou a péûÙstupnou metodu, jak zûÙskat dé₤leéƒitûÀ data o migraci. Krouéƒkuje se pomocûÙ kovovû§ch krouéƒké₤ (nejáastáji hlinûÙkovû§ch), kterûˋ se umûÙstûÙ na báhûÀk ptûÀka. PtûÀci ke krouéƒkovûÀnûÙ se nejáastáji chytajûÙ v hnûÙzdiéÀtûÙch nebo za letu do ptaáûÙch sûÙtûÙ.[175]
I kdyéƒ krouéƒkovûÀnûÙ péineslo odpovádi na éadu zûÀkladnûÙch otûÀzek spojenû§ch s migracûÙ, neposkytuje informace jako napé. kde se jedinec pohyboval mezi mûÙstem, kde byl okrouéƒkovûÀn, a odchytovû§m mûÙstem. DaléÀûÙm problûˋm krouéƒkovûÀnûÙ je nûÙzkûÀ nûÀvratnost krouéƒké₤, kterûˋ se mé₤éƒe pohybovat jen v desetinûÀch procent. Pré₤lom nastal v 90. letech 20. stoletûÙ s péûÙchodem telemetrickû§ch zaéûÙzenûÙ jako jsou satelitnûÙ vysûÙlaáe.[176]
Obecná platûÙ, éƒe telemetrickûˋ zaéûÙzenûÙ umûÙstánûˋ na ptûÀkovi by nemálo péesahovat 3 % jeho hmotnosti.[177] U velkû§ch ptûÀké₤ je moéƒnûˋ pouéƒûÙvat pokroáilûÀ a pomárná táéƒkûÀ zaéûÙzenûÙ vybavenûÀ solûÀrnûÙmi panely péijûÙmajûÙcûÙmi signûÀl satelitnûÙ navigace. Tato zaéûÙzenûÙ se mohou napéûÙklad péipevéovat jako obojky na krky nebo na rû§dovacûÙ pera.[175] Lety velkû§ch ptûÀké₤ tak lze sledovat i v reûÀlnûˋm áase.[178] U menéÀûÙch druhé₤ se pouéƒûÙvajûÙ hlavná ultralehkûˋ zaéûÙzenûÙ zvanûˋ geolokûÀtory. Tato miniaturnûÙ zaéûÙzenûÙ mohou vûÀéƒit i mûˋná neéƒ 1 gram a unesou je i tak malûˋ druhy jako vlaéÀtovky. ZaéûÙzenûÙ zûÙskûÀvûÀ v uráenûˋm áasovûˋm intervalu kaéƒdodennûÙ data o intenzitá osvátlenûÙ, z áehoéƒ se dajûÙ vyvodit zemápisnûˋ souéadnice. NejvátéÀûÙ nevû§hody geolokûÀtoré₤ jsou nutnost odchycenûÙ jedincé₤ pro stûÀhnutûÙ dat[176] a zemápisnûˋ souéadnice odvozenûˋ z geolokûÀtoré₤ majûÙ rozptyl ôÝ150 km u zemápisnûˋ éÀûÙéky a ôÝ200 km u zemápisnûˋ dûˋlky.[179] V roce 2015 doéÀlo k prvnûÙmu testu miniaturnûÙch zaéûÙzenûÙ, kterûÀ dokûÀéƒûÙ komunikovat se satelitem v uráenû§ch intervalech (v tomto péûÙpadá kaéƒdû§ 28 dnûÙ) a péesto vûÀéƒûÙ kolem 1 g. Tato zaéûÙzenûÙ byla péipevnána na hébety leséûÀáké₤ zlatohlavû§ch (vûÀha jedince kolem 20g). I kdyéƒ bylo ptûÀky nutnûˋ znovu odchytit pro stûÀhnutûÙ dat, zaéûÙzenûÙ poskytla cennûˋ informace o zimoviéÀtûÙch druhu.[179]
K daléÀûÙm metodûÀm studia migrace patéûÙ radarovûÀ zaéûÙzenûÙ[180] nebo satelitnûÙ snûÙmky.[181] Migraci ptûÀké₤ pomûÀhajûÙ studovat dokonce i astronauti z MezinûÀrodnûÙ vesmûÙrnûˋ stanice.[182] Vedle modernûÙch technologiûÙ vádci pouéƒûÙvajûÙ i mnohem jednoduéÀéÀûÙ zaéûÙzenûÙ jako je Emlené₤v trychtû§é. JednûÀ se v podstatá o plechovou klec ve tvaru péevrûÀcenûˋho kuéƒelu s malou ploéÀinkou na dná potéenou inkoustem. V dobá migrace odchycenû§ ptûÀk hopkûÀ po inkoustu a po stánûÀch plechovûˋ klece obalenûˋ papûÙrem, áûÙméƒ se pak dûÀ rozpoznat, jakû§m smárem by se nejradáji vydal. StropnûÙ áûÀst bû§vûÀ bû§t zakryta jen drûÀtánû§m pletivem, dûÙky áemuéƒ mé₤éƒe ptûÀk pozorovat, co je nad nûÙm. V péûÙpadá potéeby tak lze napé. provûÀdát pokusy v planetûÀriu pomocûÙ manipulace postavenûÙ hvázd.[183]
DaléÀûÙ modernûÙ metodou studia migrace ptûÀké₤ je rozbor stabilnûÙch izotopé₤ vodûÙké₤ (deuterium) ve tkûÀnûÙch ptûÀké₤ (napé. v ké₤éƒi nebo peéûÙ). Izotopovûˋ sloéƒenûÙ ptaáûÙch tkûÀnûÙ odrûÀéƒûÙ izotopovûˋ sloéƒenûÙ jejich potravy, kterûÀ byla hmyzem ái rostliny nasûÀknuta skrze deéÀéËovou vodu. Obsah deuteria v deéÀéËovûˋ vodá je jedineánû§ pro kaéƒdou oblast, takéƒe jeho analû§zou lze uráit mûÙsto pé₤vodu ptûÀké₤.[184][185]
OhroéƒenûÙ a ochrana
Vádeckû§ vû§zkum patéûÙ ke klûÙáovû§m metodûÀm ochrany taéƒnû§ch ptûÀké₤ (na obrûÀzku vû§zkumnûÙk dréƒûÙcûÙ leséûÀáka tajgovûˋho, Minnesota, USA)
NejvátéÀûÙ ohroéƒenûÙ stáhovavû§ch ptûÀké₤ péedstavuje álovák a jeho aktivity. Tahovûˋ cesty áasto vedou napéûÙá stûÀty, takéƒe management a ochrana ptûÀké₤ vyéƒaduje mezinûÀrodnûÙ spoluprûÀci. Pro tyto û¤áely bylo vytvoéeno nákolik mezinûÀrodnûÙch dohod. K tám nejdé₤leéƒitáéÀûÙm patéûÙ tzv. BonnskûÀ û¤mluva,[149] v rûÀmci kterûˋ byla sjednûÀna i Dohoda o ochraná africko-euroasijskû§ch stáhovavû§ch vodnûÙch ptûÀké₤ (AEWA).[186] K signatûÀéé₤m obou dohod patéûÙ i EvropskûÀ unie, potaéƒmo áesko.[149][186]
Dlouhou dobu se málo za to, éƒe stáhovavûÙ ptûÀci báhem migrace zastavujûÙ tam, kde se naskytne vhodnûÀ péûÙleéƒitost. ModernûÙ vû§zkumy se sledovacûÙmi zaéûÙzenûÙmi nicmûˋná ukûÀzaly opak; stáhovavûÙ ptûÀci áasto dálajûÙ pouze nákolik mûÀlo tahovû§ch zastûÀvek na svûˋ migraánûÙ cestá a jsou tedy citelná zûÀvislûÙ na svû§ch odpoáinkovû§ch stanoviéÀtûÙch. To platûÙ zejmûˋna v mûÙstech, kterûÀ fungujûÙ jako péirozenû§ trychtû§é migraánûÙch cest jako je napé. StéednûÙ Amerika, kterûÀ péedstavuje vzhledem k severoamerickûˋ pevniná jen û¤zkû§ pûÀs pevniny, kterû§ éada migraánûÙch ptûÀké₤ vyuéƒûÙvûÀ.[51] Tyto oblasti je tedy potéeba chrûÀnit, coéƒ je nákdy problematickûˋ, protoéƒe ptûÀci svûÀ odpoáinkovûÀ stanoviéÀtá áasto vyuéƒûÙvajûÙ jen nákolik dnûÙ v roce, a mé₤éƒe bû§t sloéƒitûˋ tato mûÙsta identifikovat.[187] Vedle ochrany stanoviéÀéË se snahy ochranûÀéé₤ soustéeáujûÙ mj. na potlaáenûÙ pytlûÀctvûÙ, omezenûÙ hazardnûÙch bariûˋr (napé. drûÀté₤ elektrickûˋho vedenûÙ, do kterû§ch taéƒnûÙ ptûÀci nákdy narûÀéƒûÙ), vádeckû§ vû§zkum nebo vzdálûÀvûÀnûÙ veéejnosti o problematice ochrany migraánûÙch druhé₤.[188]
MladûÙ jeéûÀbi ameriátûÙ nûÀsledujûÙ ultralehkûˋ letadlo báhem svûˋ prvnûÙ migrace z Wisconsinu na Floridu (leden 2009)
Pomárná báéƒnû§m zpé₤sobem ochrany ptaáûÙch druhé₤ je pomocûÙ metody reintrodukce (tzn. znovu uvedenûÙ druhu do mûÙst, odkud byl v minulosti vyhuben). U taéƒnû§ch druhé₤ to je nicmûˋná problematickûˋ ã jakmile totiéƒ dojde k vyhubenûÙ uráitûˋ populace, dojde i k zûÀniku gené₤ s informacemi o tahovûˋ cestá. ZnûÀmû§ je péûÙpad jeéûÀbé₤ americkû§ch, kteéûÙ byli kdysi rozéÀûÙéeni po celûˋm stéedozûÀpadá Spojenû§ch stûÀté₤, avéÀak nûÀsledkem lovu a drastickûˋho û¤bytku péirozenûˋho prostéedûÙ se vyskytujûÙ jiéƒ jen v nákolika mûÀlo oblastech. V rûÀmci jednoho z reintrodukánûÙch programé₤ byla do Wisconsinu (sever Spojenû§ch stûÀté₤ americkû§ch) reintrodukovûÀna malûÀ populace jeéûÀbé₤, kterûÀ byla jiéƒ od maliáka uáena nûÀsledovat ultralehkûˋ letadlo. Toto letadlo nakonec jeéûÀby vedlo pées celûˋ Spojenûˋ stûÀty aéƒ do jejich zimoviéÀéË na Floridá (jih Spojenû§ch stûÀté₤ americkû§ch). Po skonáenûÙ zimy se jeéûÀbi dovedli sami vrûÀtit zpát do Wisconsinu.[189]
Nákterûˋ druhy stáhovavû§ch ptûÀké₤ jsou nûÀsledkem pé₤sobenûÙ álováka jiéƒ vyhynulûˋ. JednûÙm z nejznûÀmájéÀûÙch péûÙpadé₤ je holub stáhovavû§, jehoéƒ poáty na severoamerickûˋm kontinentu péed péûÙchodem Evropané₤ dosahovaly kolem 3ã5 miliard. Nadmárnû§ lov, odleséovûÀnûÙ a dramatickû§ û¤bytek biotopu véÀak vedlo k vyhnutûÙ druhu.[190] Mezi stáhovavû§mi druhy je velkûˋ mnoéƒstvûÙ ohroéƒenû§ch i kriticky ohroéƒenû§ch druhé₤; velkûÀ áûÀst z nich nûÀleéƒûÙ k trubkonosû§m.[191]
Na podporu éÀûÙéenûÙ povádomûÙ o potéebá ochrany stáhovavû§ch druhé₤ se kaéƒdoroáná konûÀ Svátovû§ den stáhovavû§ch ptûÀké₤.[192]
Zmána klimatu
ProbûÙhajûÙcûÙ zmána klimatu majûÙ na migraci ptûÀké₤ dalekosûÀhlû§ vliv. DochûÀzûÙ ke zmánûÀm v ptaáûÙ fenologii, morfologii i naáasovûÀnûÙ migrace. NûÀsledkem globûÀlnûÙho oteplovûÀnûÙ nastûÀvûÀ po celûˋm svátá k áasnájéÀûÙm odleté₤m ptûÀké₤ do zimoviéÀéË, áûÙméƒ se ptûÀci snaéƒûÙ péizpé₤sobit stûÀle ranájéÀûÙmu péûÙchodu jara, se kterû§m je spojovûÀno mnoéƒstvûÙ dostupnûˋ potravy jako je nektar, ovoce a hmyz. V pré₤máru se doba odletu migranté₤ posouvûÀ od pozdnûÙho 20. stoletûÙ o pé₤l dne aéƒ nákolik dnûÙ vpéed kaéƒdû§ch deset let.[193][194][195][196]
VlaéÀtovka stromovûÀ patéûÙ k jednomu z mnoha druhé₤, u kterû§ch doéÀlo k posunu doby zahnûÙzdánûÙ vpéed
Tyto zmány naáasovûÀnûÙ véÀak mohou vûˋst k prudkû§m populaánûÙm poklesé₤m taéƒnû§ch druhé₤. PtûÀci mohou napé. péiletát do zimoviéÀéË jeéÀtá péed tûÙm, neéƒ dojde k rojenûÙ hmyzu, coéƒ vzhledem k vyáerpûÀnûÙ ptûÀké₤ po dlouhûˋm letu mé₤éƒe mûÙt kritickûˋ nûÀsledky.[196] V dé₤sledku zmány naáasovûÀnûÙ dochûÀzûÙ i ke zmánûÀm v hnûÙzdnûÙch zvyklostech stáhovavû§ch ptûÀké₤.[194] Napé. doba hnûÙzdánûÙ severoamerickûˋ vlaéÀtovky stromovûˋ se jen mezi lety 1959ã1991 posunula vpéed v pré₤máru o 9 dnûÙ, coéƒ je vedle vyéÀéÀûÙch teplot spojovûÀno i s ranájéÀûÙ dobou kvátenstvûÙ tamájéÀûÙ vegetace.[197] U nizozemskû§ch populacûÙ lejsáûÙké₤ áernohlavû§ch sice doéÀlo k posunutûÙ doby kladenûÙ vajec vpéed, takéƒe ptûÀci majûÙ dostatek potravy v dobá hnûÙzdánûÙ, nicmûˋná nedoéÀlo k posunutûÙ samotnûˋho migraánûÙho letu. PtûÀci tak péilûˋtajûÙ do NizozemûÙ péûÙliéÀ pozdá na to, aby stihli vrcholnûˋ obdobûÙ vû§skytu housenek, kterûˋ péedstavujûÙ jeden z jejich klûÙáovû§ch potravnûÙch zdrojé₤. NûÀsledkem toho nákterûˋ nizozemskûˋ populace lejsáûÙké₤ áernohlavû§ch poklesly mezi lety péibliéƒná 1985ã1995 aéƒ o 90 %.[198] Podle vû§sledké₤ studie 20 druhé₤ evropskû§ch migraánûÙch druhé₤ se pré₤márnûÀ doba migranté₤ strûÀvenûÀ v Evropá mezi lety 1992ã2018 prodlouéƒila o 16 dnûÙ. Autoéi studie péedpoklûÀdajûÙ, éƒe nákterûˋ druhy jako konipasi, slavûÙk obecnû§ ái budnûÙáek vátéÀûÙ mohou û¤plná péestat migrovat.[199][200]
U éady druhé₤ dochûÀzûÙ ke zkracovûÀnûÙ dûˋlky migrace, protoéƒe ptûÀci jiéƒ nemusûÙ lûˋtat tak daleko, aby naéÀli vhodnûˋ podmûÙnky pro péezimovûÀnûÙ.[201] PéûÙkladem z áeskûˋho ptactva mohou bû§t áûÀpi bûÙlûÙ, kteéûÙ nákdy hnûÙzdûÙ v éecku nebo é panálsku mûÙsto v Africe, nebo nákterûˋ populace pánic áernohlavû§ch, kterûˋ zimujûÙ ve Velkûˋ BritûÀnii namûÙsto v Africe.[143] S vyéÀéÀûÙmi teplotami jsou spojovûÀny i zmány v morfologii ptûÀké₤ ã u migrujûÙcûÙch druhé₤ dochûÀzûÙ k postupnûˋmu prodluéƒovûÀnûÙ kéûÙdel a zmenéÀovûÀnûÙ tál, coéƒ je spojovûÀno s efektivnájéÀûÙ migracûÙ. MenéÀûÙ tála totiéƒ lûˋpe odvûÀdûÙ teplo z rozehéûÀtû§ch tál letûÙcûÙch ptûÀké₤ a deléÀûÙ kéûÙdla zefektivéujûÙ lûˋtûÀnûÙ.[202][203]
NejslibnájéÀûÙ obranou ptûÀké₤ proti zmánûÀm klimatu je flexibilita migrace. áetnûˋ vû§zkumy ukazujûÙ, éƒe û¤prava naáasovûÀnûÙ migrace vzhledem ke klimatickû§m zmánûÀm mûÀ pozitivnûÙ vliv na populaci druhu.[204] ZdûÀ se, éƒe migranti na krûÀtkou vzdûÀlenost dokûÀéƒûÙ reagovat na zmány klimatu pruéƒnáji neéƒ migranti na dlouhou vzdûÀlenost. DûÀlkovûÙ migranti totiéƒ áasujûÙ své₤j tah hlavná na zûÀkladá mnoéƒstvûÙ dennûÙho svátla a nemohou na dûÀlku odhadnout podmûÙnky destinace. Naproti tomu migranti tûÀhnoucûÙ krûÀtkûˋ vzdûÀlenosti mohou lûˋpe vytuéÀit podmûÙnky v zimoviéÀti / hnûÙzdiéÀti a mohou tak dokonce profitovat z probûÙhajûÙcûÙch zmán klimatu (napé. v péûÙpadá áasnûˋho oteplenûÙ hnûÙzdiéÀtá se tam mohou vypravit déûÙve, coéƒ jim mé₤éƒe poskytnout konkurenánûÙ vû§hodu).[205] Nicmûˋná û¤prava naáasovûÀnûÙ migrace je u kaéƒdûˋho druhu jinûÀ a ne véƒdy se dáje dostateáná rychle nebo pruéƒná na to, aby se vyrovnala s nûÀsledky klimatickû§ch zmán.[196]
Odraz v kultuée
MigraánûÙ druhy ptûÀké₤ zanechaly svou stopu v mnohû§ch kulturûÀch v podobá mû§té₤, legend ái tradiánûÙch domorodû§ch tancé₤.[206] Ve Starovákûˋm Egyptá bylo na kaéƒdoroánûÙ péûÙlet a odlet migraánûÙch druhé₤ nahlûÙéƒeno jako na souáûÀst cyklickûˋho bytûÙ sváta.[207] Ve staroáûÙnskûˋ sbûÙrce texté₤ Chuaj-nan-c' je kaéƒdoroánûÙ odlet ptûÀké₤ péiáûÙtûÀn na vrub metamorfû°ze, áili péemáná ptaáûÙch druhé₤ na jinûˋ bytosti. Ve sbûÙrce se mj. pûÙéÀe, éƒe vlaéÀtovky na podzim nalûˋtajûÙ do moée a péemánûÙ se ve éÀkeble. Podobná i podle staréÀûÙch áûÙnskû§ch kalendûÀéé₤ se jestéûÀbi na jaée mánûÙ v holuby a myéÀi v kéepelky.[207] Migrace ptûÀké₤ je áasto spojovûÀna se zmánou roánûÙch dob.[206] Napé. v áesku se vlaéÀtovky a jiéiáky péezdûÙvajûÙ ãposlovûˋ jaraã.[208] Nákdy se za posly jara oznaáujûÙ i áûÀpi bûÙlûÙ,[209] kteéûÙ jsou hojná spojovûÀni i s noéÀenûÙm dátûÙ.[210] NoéÀenûÙ dátûÙ áûÀpy zváánil dûÀnskû§ klasik Hans Christian Andersen ve svûˋ ponákud potemnálûˋ pohûÀdce ãáûÀpiã. é vûˋdskûÀ spisovatelka Selma LagerlûÑf v knize PodivuhodnûÀ cesta Nilse Holgerssona é vûˋdskem popsala péûÙbáh chlapce Nilse, kterû§ cestuje é vûˋdskem na hébetu husy.[211]
StáhovûÀnûÙ ptûÀké₤ se stalo nûÀmátem éady dokumentûÀrnûÙch filmé₤ jako je napé. PtaáûÙ svát[212] nebo nákolika dûÙlé₤ dokumentûÀrnûÙ minisûˋrie Migrations secrû´tes (ZûÀhady migrace).[213] O zmûÙnánûˋm vedenûÙ hus a jeéûÀbé₤ ultralehkû§m letadlem byl natoáen film Cesta domé₤ (1996).[214]
ãROLLAND, Jonathan; JIGUET, Frûˋdûˋric; JûNSSON, Knud Andreas; CONDAMINE, Fabien L.; MORLON, Hûˋlû´ne. Settling down of seasonal migrants promotes bird diversification. S. 20140473. Proceedings of the Royal Society B: Biological Sciences [online]. 2014-06-07 [cit. 2021-12-11]. Roá. 281, áûÙs. 1784, s. 20140473. DOI10.1098/rspb.2014.0473. (anglicky)
ã abSOMVEILLE, Marius; MANICA, Andrea; BUTCHART, Stuart H. M.; RODRIGUES, Ana S. L. Mapping Global Diversity Patterns for Migratory Birds. S. e70907. PLoS ONE [online]. 2013-08-07 [cit. 2021-12-20]. Roá. 8, áûÙs. 8, s. e70907. DOI10.1371/journal.pone.0070907. (anglicky)
ã abThe Basics of Bird Migration: How, Why, and Where [online]. The Cornell Lab, 2021-08-01 [cit. 2021-12-11]. Dostupnûˋ online. (anglicky)
ã abcBOERE, Gerard C.; STROUD, David A. The flyway concept: what it is and what it isnãt. In: BOERE, G. C.; GALBRAITH, C. A.; STROUD, D. A. Waterbirds around the world: a global overview of the conservation, management and research of the world's waterbird flyways. Edinburgh: The Stationery Office, 2006. Dostupnûˋ online. ISBN0114973334. (anglicky)
ãPANOV, Ilya N. Overlap between moult and autumn migration in passerines in northern taiga zone of Eastern Fennoscandia. S. 33ã64. Avian Ecology and Behaviour [online]. 2011 [cit. 2021-12-12]. Roá. 19, s. 33ã64. Dostupnûˋ online. (anglicky)
ãMEISSNER, Wéodzimierz. Immature dunlins Calidris alpina migrate towards wintering grounds later than adults in years of low breeding success. S. 47ã53. Journal of Ornithology [online]. 2015-01 [cit. 2021-12-12]. Roá. 156, áûÙs. 1, s. 47ã53. DOI10.1007/s10336-014-1132-y. (anglicky)
ãMEISSNER, Wéodzimierz; WéODARCZAK-KOMOSIéSKA, Anna; GûRECKI, Dariusz; WûJCIK, Cezary; éCIBORSKI, Mateusz; KRUPA, Robert; ZIáCIK, Piotr. Autumn Migration of Waders (Charadrii) at the Reda Mouth (N Poland). S. 23ã39. RING [online]. 2009-01-01. Roá. 31, áûÙs. 1, s. 23ã39. DOI10.2478/v10050-008-0046-6. (anglicky)
ãKETTERSON, E. D.; NOLAN, V. Intraspecific variation in avian migration: evolutionary and regulatory aspects. In: RANKIN, M. A. Migration: mechanisms and adaptive significance. University of Texas, Austin: University of Texas Press, 1985. S. 553ã579.
ãSCOTT, Graham R.; HAWKES, Lucy A.; FRAPPELL, Peter B.; BUTLER, Patrick J.; BISHOP, Charles M.; MILSOM, William K. How Bar-Headed Geese Fly Over the Himalayas. S. 107ã115. Physiology [online]. 2015-03 [cit. 2021-12-14]. Roá. 30, áûÙs. 2, s. 107ã115. DOI10.1152/physiol.00050.2014. (anglicky)
ãHAWKES, L. A.; BALACHANDRAN, S.; BATBAYAR, N.; BUTLER, P. J.; FRAPPELL, P. B.; MILSOM, W. K.; TSEVEENMYADAG, N. The trans-Himalayan flights of bar-headed geese (Anser indicus). S. 9516ã9519. Proceedings of the National Academy of Sciences [online]. 2011-06-07 [cit. 2021-12-14]. Roá. 108, áûÙs. 23, s. 9516ã9519. DOI10.1073/pnas.1017295108. (anglicky)
ãSENNER, Nathan R.; STAGER, Maria; VERHOEVEN, Mo A.; CHEVIRON, Zachary A.; PIERSMA, Theunis; BOUTEN, Willem. High-altitude shorebird migration in the absence of topographical barriers: avoiding high air temperatures and searching for profitable winds. S. 20180569. Proceedings of the Royal Society B: Biological Sciences [online]. 2018-06-27 [cit. 2021-12-14]. Roá. 285, áûÙs. 1881, s. 20180569. DOI10.1098/rspb.2018.0569. (anglicky)
ãCORSO, Andrea. European Birding Hot Spot: The Strait of Messina, southern Italy. [online]. Surfbirds [cit. 2021-12-14]. Dostupnûˋ online. (anglicky)
ãKolektiv autoré₤. Strategie ochrany biologickûˋ rozmanitosti áeskûˋ republiky 2016ã2025. [s.l.]: Ministerstvo éƒivotnûÙho prostéedûÙ, 2016. Dostupnûˋ v archivu poéûÙzenûˋm dne 2022-01-19. ISBN978-80-7212-609-5. S. 112.
ãRUEGG, Kristen C.; SMITH, Thomas B. Not as the crow flies: a historical explanation for circuitous migration in Swainson's thrush (Catharus ustulatus). S. 1375ã1381. Proceedings of the Royal Society of London. Series B: Biological Sciences [online]. 2002-07-07 [cit. 2021-12-13]. Roá. 269, áûÙs. 1498, s. 1375ã1381. DOI10.1098/rspb.2002.2032. (anglicky)
ãKLVAéA, Petr. PodzimnûÙ cesty naéÀich taéƒnû§ch ptûÀké₤. S. 11ã15. PtaáûÙ svát [online]. áeskûÀ spoleánost ornitologickûÀ, 2017 [cit. 2021-12-16]. Roá. XXIV, áûÙs. 3, s. 11ã15.
ãRAMACHANDRA, T. V.; MAHAPATRA, Durga Madhab; RAO, Sankara Kolluri; NATHAN, M. Bhoomi. Environmental Impact Assessment of the National Large Solar Telescope Project and Its Ecological Impact in Merak Area [online]. Energy & Wetland Research Group Centre for Ecological Sciences, Indian Institute of Science, Bengalore, India, 2013 [cit. 2021-12-11]. S. 69. Dostupnûˋ online. (anglicky)
ãJAHN, Alex E.; BRAVO, Susana P.; CUETO, VûÙctor R. Patterns of partial avian migration in northern and southern temperate latitudes of the New World. Emu - Austral Ornithology. 2012-03-01, roá. 112, áûÙs. 1, s. 17ã22. Dostupnûˋ online [cit. 2021-12-12]. ISSN0158-4197. DOI10.1071/MU10091. (anglicky)
ãWhat We Do [online]. Birdlife International [cit. 2021-12-14]. Dostupnûˋ online. (anglicky)
ã ab Stopover Habitats and What Birds Need on Migration. American Bird Conservancy [online]. 2017-04-27 [cit. 2021-12-14]. Dostupnûˋ online. (anglicky)
ãFERRETTI, Andrea; MAGGINI, Ivan; LUPI, Sara. The amount of available food affects diurnal locomotor activity in migratory songbirds during stopover. Scientific Reports. 2019-12, roá. 9, áûÙs. 1, s. 19027. Dostupnûˋ online [cit. 2021-12-14]. ISSN2045-2322. DOI10.1038/s41598-019-55404-3. PMID31836848. (anglicky)
ãVrcholûÙ migrace ptûÀké₤ ã pomozme jim v letu o éƒivot! ã áeskûÀ spoleánost ornitologickûÀVrcholûÙ migrace ptûÀké₤ ã pomozme jim v letu o éƒivot! [online]. áeskûÀ spoleánost ornitologickûÀ, 2021-10-19 [cit. 2021-12-14]. Dostupnûˋ online. (anglicky)
ã The ecology, behavior, and conservation of migratory birds. www.usgs.gov [online]. U.S. Geological Survey [cit. 2022-01-26]. Dostupnûˋ online. (anglicky)
ãNUWER, Rachel. Fish Eggs Survive Journey through a Duck [online]. Scientific American, 2020-09-01 [cit. 2022-01-26]. Dostupnûˋ online. (anglicky)
ãLENNOX, Robert J.; CHAPMAN, Jacqueline M.; SOULIERE, Christopher M.; TUDORACHE, Christian; WIKELSKI, Martin; METCALFE, Julian D.; COOKE, Steven J. Conservation physiology of animal migration. Conservation Physiology [online]. 2016 [cit. 2022-01-26]. Roá. 4, áûÙs. 1. DOI10.1093/conphys/cov072. (anglicky)
ãFRITZSCHE MCKAY, Alexa; HOYE, Bethany J. Are Migratory Animals Superspreaders of Infection?. S. 260ã267. Integrative and Comparative Biology [online]. 2016-08-01 [cit. 2022-01-25]. Roá. 56, áûÙs. 2, s. 260ã267. DOI10.1093/icb/icw054. (anglicky)
ãCOHEN, Emily B.; AUCKLAND, Lisa D.; MARRA, Peter P.; HAMER, Sarah A. Avian Migrants Facilitate Invasions of Neotropical Ticks and Tick-Borne Pathogens into the United States. S. 8366ã8378. Applied and Environmental Microbiology [online]. 2015-12-15 [cit. 2022-01-25]. Roá. 81, áûÙs. 24, s. 8366ã8378. DOI10.1128/AEM.02656-15. (anglicky)
ãVERHAGEN, Josanne H.; HERFST, Sander; FOUCHIER, Ron A. M. How a virus travels the world. S. 616ã617. Science [online]. 2015-02-06 [cit. 2022-01-25]. Roá. 347, áûÙs. 6222, s. 616ã617. DOI10.1126/science.aaa6724. (anglicky)
ãTURNER, Angela K. Swallows & martins: an identification guide and handbook. Boston: Houghton Mifflin, 1989. Dostupnûˋ online. ISBN0395511747. S. 164ã169. (anglicky)
ãVAN GILS, Jan A; BATTLEY, Phil F; PIERSMA, Theunis; DRENT, Rudi. Reinterpretation of gizzard sizes of red knots world-wide emphasises overriding importance of prey quality at migratory stopover sites. S. 2609ã2618. Proceedings of the Royal Society B: Biological Sciences [online]. 2005-12-22 [cit. 2021-12-14]. Roá. 272, áûÙs. 1581, s. 2609ã2618. DOI10.1098/rspb.2005.3245. (anglicky)
ãLongest non-stop migration by a bird [online]. Guinness World Records [cit. 2021-04-30]. Dostupnûˋ online. (anglicky)
ãHROMûDKOVû, T; PAVEL, V; FLOUSEK, J; BRIEDIS, M. Seasonally specific responses to wind patterns and ocean productivity facilitate the longest animal migration on Earth. S. 1ã12. Marine Ecology Progress Series [online]. 2020-03-19 [cit. 2021-12-11]. Roá. 638, s. 1ã12. DOI10.3354/meps13274. (anglicky)
ãáûÀpi a vlaéÀtovky se s tûÙm moc nepûÀéou, do Afriky doletûÙ na jeden zûÀtah [online]. áeskû§ rozhlas, 2017-10-17 [cit. 2021-12-20]. Dostupnûˋ online.
ãBAIRLEIN, Franz; NORRIS, D. Ryan; NAGEL, Rolf; BULTE, Marc; VOIGT, Christian C.; FOX, James W.; HUSSELL, David J. T. Cross-hemisphere migration of a 25 g songbird. S. 505ã507. Biology Letters [online]. 2012-08-23. Roá. 8, áûÙs. 4, s. 505ã507. DOI10.1098/rsbl.2011.1223. (anglicky)
ãFeatherweight songbird is a long-distance champ [online]. Phys.org [cit. 2021-12-14]. Dostupnûˋ online. (anglicky)
ãSHAFFER, S. A.; TREMBLAY, Y.; WEIMERSKIRCH, H.; SCOTT, D.; THOMPSON, D. R.; SAGAR, P. M.; MOLLER, H. Migratory shearwaters integrate oceanic resources across the Pacific Ocean in an endless summer. S. 12799ã12802. Proceedings of the National Academy of Sciences [online]. 2006-08-22 [cit. 2021-05-18]. Roá. 103, áûÙs. 34, s. 12799ã12802. DOI10.1073/pnas.0603715103. (anglicky)
ãBARûANTE, Luciana; M. VALE, Mariana; S. ALVES, Maria Alice. Altitudinal migration by birds: a review of the literature and a comprehensive list of species. S. 321ã335. Journal of Field Ornithology [online]. 2017-12 [cit. 2021-12-16]. Roá. 88, áûÙs. 4, s. 321ã335. DOI10.1111/jofo.12234. (anglicky)
ãBOYLE, W. Alice. Altitudinal bird migration in North America. S. 443ã465. The Auk [online]. 2017-04 [cit. 2021-12-16]. Roá. 134, áûÙs. 2, s. 443ã465. DOI10.1642/AUK-16-228.1. (anglicky)
ãBOYLE, W. Alice; CONWAY, Courtney J.; BRONSTEIN, Judith L. Why do some, but not all, tropical birds migrate? A comparative study of diet breadth and fruit preference. S. 219ã236. Evolutionary Ecology [online]. 2011-01 [cit. 2021-12-16]. Roá. 25, áûÙs. 1, s. 219ã236. DOI10.1007/s10682-010-9403-4. (anglicky)
ãHOBSON, Keith A.; WASSENAAR, Len I.; MILû, Borja; LOVETTE, Irby; DINGLE, Caroline; SMITH, Thomas B. Stable isotopes as indicators of altitudinal distributions and movements in an Ecuadorean hummingbird community. S. 302ã308. Oecologia [online]. 2003-07-01 [cit. 2021-12-16]. Roá. 136, áûÙs. 2, s. 302ã308. DOI10.1007/s00442-003-1271-y. (anglicky)
ãShortest bird migration [online]. Guinness World Records [cit. 2021-12-16]. Dostupnûˋ online. (anglicky)
ãCHAN, Ken. Partial migration in Australian landbirds: a review. S. 281ã292. Emu - Austral Ornithology [online]. 2001-12 [cit. 2021-12-12]. Roá. 101, áûÙs. 4, s. 281ã292. DOI10.1071/MU00034. (anglicky)
ãDINGLE, Hugh. Bird migration in the southern hemisphere: a review comparing continents. S. 341ã359. Emu - Austral Ornithology [online]. 2008-12 [cit. 2021-12-12]. Roá. 108, áûÙs. 4, s. 341ã359. DOI10.1071/MU08010. (anglicky)
ãLUNDBERG, Per. The evolution of partial migration in Birds. S. 172ã175. Trends in Ecology & Evolution [online]. 1988-07 [cit. 2021-12-12]. Roá. 3, áûÙs. 7, s. 172ã175. DOI10.1016/0169-5347(88)90035-3. (anglicky)
ãBOLAND, John M. Leapfrog Migration in North American Shorebirds: Intra- and Interspecific Examples. The Condor [online]. Oxford University Press, 1990-05 [cit. 2021-12-12]. Roá. 92, áûÙs. 2. DOI10.2307/1368226. (anglicky)
ãBELL, Christopher P. Leap-Frog Migration in the Fox Sparrow: Minimizing the Cost of Spring Migration. S. 470ã477. The Condor [online]. 1997-05 [cit. 2021-12-12]. Roá. 99, áûÙs. 2, s. 470ã477. DOI10.2307/1369953. (anglicky)
ãû KESSON, Susanne; ATKINSON, Phil W.; BERMEJO, Ana; PUENTE, Javier; FERRI, Mauro; HEWSON, Chris M.; HOLMGREN, Jan. Evolution of chain migration in an aerial insectivorous bird, the common swift Apus apus. S. 2377ã2391. Evolution [online]. 2020-10 [cit. 2021-12-12]. Roá. 74, áûÙs. 10, s. 2377ã2391. DOI10.1111/evo.14093. (anglicky)
ãALERSTAM, Thomas. Flight by night or day? Optimal daily timing of bird migration. Journal of Theoretical Biology. 2009-06, roá. 258, áûÙs. 4, s. 530ã536. Dostupnûˋ online [cit. 2021-12-16]. DOI10.1016/j.jtbi.2009.01.020. (anglicky)
ãEleonora's Falcons imprison live birds for a later meal [online]. 2015-08-26 [cit. 2021-12-11]. Dostupnûˋ online. (anglicky)
ãQNINBA, A.; BENHOUSSA, A.; RADI, M.; EL IDRISSI, A.; BOUSADIK, H.; BADAOUI, B.; EL AGBANI, M. A. Mode de prûˋdation trû´s particulier du Faucon dãûlûˋonore (Falco eleonorae) sur lãArchipel dãEssaouira, Maroc Atlantique. S. 149ã150. Alauda [online]. 2015 [cit. 2021-12-11]. Roá. 83, s. 149ã150. (francouzsky)
ãIBANEZ, C.; JUSTE, J.; GARCIA-MUDARRA, J. L.; AGIRRE-MENDI, P. T. Bat predation on nocturnally migrating birds. S. 9700ã9702. Proceedings of the National Academy of Sciences [online]. 2001-08-14 [cit. 2021-12-11]. Roá. 98, áûÙs. 17, s. 9700ã9702. DOI10.1073/pnas.171140598. (anglicky)
ã abcRATTENBORG, Niels C; MANDT, Bruce H; OBERMEYER, William H; WINSAUER, Peter J; HUBER, Reto; WIKELSKI, Martin; BENCA, Ruth M. Migratory Sleeplessness in the White-Crowned Sparrow (Zonotrichia leucophrys gambelii). S. e212. PLoS Biology [online]. 2004-07-13 [cit. 2021-12-20]. Roá. 2, áûÙs. 7, s. e212. DOI10.1371/journal.pbio.0020212. (anglicky)
ãFARNSWORTH, Andrew. Flight Calls and Their Value for Future Ornithological Studies and Conservation Research. S. 733ã746. The Auk [online]. 2005-07-01 [cit. 2021-12-15]. Roá. 122, áûÙs. 3, s. 733ã746. DOI10.1093/auk/122.3.733. (anglicky)
ãGRIFFITHS, Emily T.; KEEN, Sara C.; LANZONE, Michael; FARNSWORTH, Andrew. Can Nocturnal Flight Calls of the Migrating Songbird, American Redstart, Encode Sexual Dimorphism and Individual Identity?. S. e0156578. PLOS ONE [online]. 2016-06-10 [cit. 2021-12-15]. Roá. 11, áûÙs. 6, s. e0156578. DOI10.1371/journal.pone.0156578. (anglicky)
ã abPULIDO, Francisco. The Genetics and Evolution of Avian Migration. S. 165ã174. BioScience [online]. 2007-02-01 [cit. 2021-12-12]. Roá. 57, áûÙs. 2, s. 165ã174. DOI10.1641/B570211. (anglicky)
ãCHERNETSOV, Nikita; BERTHOLD, Peter; QUERNER, Ulrich. Migratory orientation of first-year white storks ( Ciconia ciconia ): inherited information and social interactions. S. 937ã943. Journal of Experimental Biology [online]. 2004-02-22 [cit. 2021-12-14]. Roá. 207, áûÙs. 6, s. 937ã943. DOI10.1242/jeb.00853. (anglicky)
ãTOEWS, David P. L.; TAYLOR, Scott A.; STREBY, Henry M.; KRAMER, Gunnar R.; LOVETTE, Irby J. Selection on VPS13A linked to migration in a songbird. S. 18272ã18274. Proceedings of the National Academy of Sciences [online]. 2019-09-10 [cit. 2021-12-20]. Roá. 116, áûÙs. 37, s. 18272ã18274. DOI10.1073/pnas.1909186116. (anglicky)
ãPULIDO, F.; BERTHOLD, P.; MOHR, G.; QUERNER, U. Heritability of the timing of autumn migration in a natural bird population. S. 953ã959. Proceedings of the Royal Society of London. Series B: Biological Sciences [online]. 2001-05-07 [cit. 2021-12-12]. Roá. 268, áûÙs. 1470, s. 953ã959. DOI10.1098/rspb.2001.1602. (anglicky)
ãBERTHOLD, P.; HELBIG, A. J.; MOHR, G.; QUERNER, U. Rapid microevolution of migratory behaviour in a wild bird species. S. 668ã670. Nature [online]. 1992-12. Roá. 360, áûÙs. 6405, s. 668ã670. DOI10.1038/360668a0. (anglicky)
ãYOUNG, Ed. On The Evolution of Migration [online]. National Geographic, 2014-08-04 [cit. 2021-12-12]. Dostupnûˋ online. (anglicky)
ãOrigin and evolution of migration [online]. Britannica [cit. 2021-12-12]. Dostupnûˋ online. (anglicky)
ã abWINGER, Benjamin M.; AUTERI, Giorgia G.; PEGAN, Teresa M.; WEEKS, Brian C. A long winter for the Red Queen: rethinking the evolution of seasonal migration. S. 737ã752. Biological Reviews [online]. 2019-06 [cit. 2021-12-13]. Roá. 94, áûÙs. 3, s. 737ã752. DOI10.1111/brv.12476. (anglicky)
ã abSALEWSKI, Volker; BRUDERER, Bruno. The evolution of bird migrationãa synthesis. S. 268ã279. Naturwissenschaften [online]. 2007-03-20 [cit. 2021-12-13]. Roá. 94, áûÙs. 4, s. 268ã279. DOI10.1007/s00114-006-0186-y. (anglicky)
ã abThe Evolution of Bird Migration [online]. The Cornell Lab, 2017-04-11 [cit. 2021-12-13]. Dostupnûˋ online. (anglicky)
ãZINK, Robert M.; GARDNER, Aubrey S. Glaciation as a migratory switch. S. e1603133. Science Advances [online]. 2017-09-29 [cit. 2021-12-13]. Roá. 3, áûÙs. 9, s. e1603133. DOI10.1126/sciadv.1603133. (anglicky)
ã abGWINNER, E. Circadian and circannual programmes in avian migration. S. 39ã48. Journal of Experimental Biology [online]. 1996-01-01 [cit. 2021-12-20]. Roá. 199, áûÙs. 1, s. 39ã48. DOI10.1242/jeb.199.1.39. (anglicky)
ãPOKROVSKY, Ivan; KûLZSCH, Andrea; SHERUB, Sherub; FIEDLER, Wolfgang; GLAZOV, Peter; KULIKOVA, Olga; WIKELSKI, Martin. Longer days enable higher diurnal activity for migratory birds. S. 2161ã2171. Journal of Animal Ecology [online]. 2021-09 [cit. 2021-12-20]. Roá. 90, áûÙs. 9, s. 2161ã2171. DOI10.1111/1365-2656.13484. (anglicky)
ãWILTSCHKO, Roswitha; WILTSCHKO, Wolfgang. Chapter Seven - Avian Navigation: A Combination of Innate and Learned Mechanisms. PéûÙprava vydûÀnûÙ Marc Naguib, H. Jane Brockmann, John C. Mitani, Leigh W. Simmons, Louise Barrett, Sue Healy, Peter J. B. Slater. Svazek 47. [s.l.]: Academic Press, 2015. Dostupnûˋ online. DOI10.1016/bs.asb.2014.12.002. S. 229ã310. (anglicky)
ãREYNOLDS, Andrew M.; CECERE, Jacopo G.; PAIVA, Vitor H. Pelagic seabird flight patterns are consistent with a reliance on olfactory maps for oceanic navigation. Proceedings of the Royal Society B: Biological Sciences. 2015-07-22, roá. 282, áûÙs. 1811, s. 20150468. Dostupnûˋ online [cit. 2021-12-22]. ISSN0962-8452. DOI10.1098/rspb.2015.0468. PMID26136443. (anglicky)
ã ab The Basics: How Birds Navigate When They Migrate. All About Birds [online]. 2008-04-01 [cit. 2021-12-22]. Dostupnûˋ online. (anglicky)
ãLûZéOVSKû, MatouéÀ. CestovûÀnûÙ podle siloáar v oku [online]. Lidovky, 2008-05-02 [cit. 2021-12-15]. Dostupnûˋ online.
ãHEYERS, Dominik; MANNS, Martina; LUKSCH, Harald; GûNTûRKûN, Onur; MOURITSEN, Henrik. A Visual Pathway Links Brain Structures Active during Magnetic Compass Orientation in Migratory Birds. S. e937. PLoS ONE [online]. 2007-09-26 [cit. 2021-12-15]. Roá. 2, áûÙs. 9, s. e937. DOI10.1371/journal.pone.0000937. (anglicky)
ã abOGDEN, Lesley Evans. Vagrant Birds May Portend Species Distribution in Climate-Changed World. Scientific American [online]. 2016-08-19 [cit. 2021-12-28]. Dostupnûˋ online. (anglicky)
ãTHORUP, Kasper; ALERSTAM, Thomas; HAKE, Mikael. Bird orientation: compensation for wind drift in migrating raptors is age dependent. Proceedings of the Royal Society of London. Series B: Biological Sciences. 2003-08-07, roá. 270, áûÙs. suppl_1, s. S8ãS11. Dostupnûˋ online [cit. 2021-12-28]. DOI10.1098/rsbl.2003.0014. PMID12952622. (anglicky)
ãELKINS, Norman. Weather and bird migration. S. 238ã256. British Birds [online]. 2005 [cit. 2021-12-28]. Roá. 98, s. 238ã256. (anglicky)
ãSANDBERG, Roland. Interaction of body condition and magnetic orientation in autumn migrating robins, Erithacus rubecula. S. 679ã686. Animal Behaviour [online]. 1994-03-01 [cit. 2021-12-28]. Roá. 47, áûÙs. 3, s. 679ã686. DOI10.1006/anbe.1994.1092. (anglicky)
ã abGILROY, James; LEES, Alexander Charles. Vagrancy theories: Are autumn vagrants really reverse migrants?. S. 427ã438. British Birds [online]. 2003 [cit. 2021-12-28]. Roá. 96, áûÙs. 9, s. 427ã438. Dostupnûˋ online. (anglicky)
ãLEES, Alexander C.; GILROY, James J. Bird migration: When vagrants become pioneers. S. R1568ãR1570. Current Biology [online]. 2021-12-20 [cit. 2021-12-28]. Roá. 31, áûÙs. 24, s. R1568ãR1570. DOI10.1016/j.cub.2021.10.058. (anglicky)
ãDUFOUR, Paul; DE FRANCESCHI, Christophe; DONIOL-VALCROZE, Paul; JIGUET, Frûˋdûˋric; GUûGUEN, Maya; RENAUD, Julien; LAVERGNE, Sûˋbastien. A new westward migration route in an Asian passerine bird. S. 5590ã5596.e4. Current Biology [online]. 2021-12. Roá. 31, áûÙs. 24, s. 5590ã5596.e4. DOI10.1016/j.cub.2021.09.086. (anglicky)
ãGUGLIELMO, Christopher G. Obese super athletes: fat-fueled migration in birds and bats. Journal of Experimental Biology. 2018-03-07, roá. 221, áûÙs. Suppl_1, s. jeb165753. Dostupnûˋ online [cit. 2021-12-27]. ISSN0022-0949. DOI10.1242/jeb.165753. (anglicky)
ã Bird Migration | How Do Birds Prepare For Migration?. The RSPB [online]. [cit. 2021-12-28]. Dostupnûˋ online. (anglicky)
ãGOYMANN, Wolfgang; LUPI, Sara; KAIYA, Hiroyuki. Ghrelin affects stopover decisions and food intake in a long-distance migrant. Proceedings of the National Academy of Sciences. 2017-02-21, roá. 114, áûÙs. 8, s. 1946ã1951. PMID 28167792. Dostupnûˋ online [cit. 2021-12-28]. ISSN0027-8424. DOI10.1073/pnas.1619565114. PMID28167792. (anglicky)
ãARAûJO, Pedro M.; VIEGAS, Ivan; ROCHA, Afonso D. Understanding how birds rebuild fat stores during migration: insights from an experimental study. Scientific Reports. 2019-07-11, roá. 9, áûÙs. 1, s. 10065. Dostupnûˋ online [cit. 2021-12-27]. ISSN2045-2322. DOI10.1038/s41598-019-46487-z. PMID31296911. (anglicky)
ã abBAUCHINGER, Ulf; KOLB, Harald; AFIK, Danny. Blackcap Warblers Maintain Digestive Efficiency by Increasing Digesta Retention Time on the First Day of Migratory Stopover. Physiological and Biochemical Zoology. 2009-09, roá. 82, áûÙs. 5, s. 541ã548. Dostupnûˋ online [cit. 2022-01-03]. ISSN1522-2152. DOI10.1086/603638. (anglicky)
ãHUME, I. D.; BIEBACH, H. Digestive tract function in the long-distance migratory garden warbler, Sylvia borin. Journal of Comparative Physiology B. 1996-10, roá. 166, áûÙs. 6, s. 388ã395. Dostupnûˋ online [cit. 2022-01-03]. ISSN0174-1578. DOI10.1007/BF02336922. (anglicky)
ãPIERSMA, Theunis; GILL, Robert E. Guts Don't Fly: Small Digestive Organs in Obese Bar-Tailed Godwits. S. 196ã203. The Auk [online]. 1998 [cit. 2021-05-01]. Roá. 115, áûÙs. 1, s. 196ã203. DOI10.2307/4089124. (anglicky)
ãBATTLEY, Phil F.; PIERSMA, Theunis. Body Composition and Flight Ranges of Bar-Tailed Godwits (Limosa lapponica baueri) from New Zealand (Composition Corporelle et Distance de Vol chez Limosa lapponica baueri de Nouvelle-Zûˋlande). S. 922ã937. The Auk [online]. 2005 [cit. 2021-05-01]. Roá. 122, áûÙs. 3, s. 922ã937. Dostupnûˋ online. ISSN0004-8038. (anglicky)
ãLANDYS-CIANNELLI, M. M.; PIERSMA, T.; JUKEMA, J. Strategic size changes of internal organs and muscle tissue in the Bar-tailed Godwit during fat storage on a spring stopover site. Functional Ecology. 2003-04, roá. 17, áûÙs. 2, s. 151ã159. Dostupnûˋ online [cit. 2022-01-03]. ISSN0269-8463. DOI10.1046/j.1365-2435.2003.00715.x. (anglicky)
ãRATTENBORG, Niels C.; VOIRIN, Bryson; CRUZ, Sebastian M. Evidence that birds sleep in mid-flight. Nature Communications. 2016-08-03, roá. 7, áûÙs. 1, s. 12468. Dostupnûˋ online [cit. 2021-12-28]. ISSN2041-1723. DOI10.1038/ncomms12468. PMID27485308. (anglicky)
ãLIECHTI, Felix; WITVLIET, Willem; WEBER, Roger. First evidence of a 200-day non-stop flight in a bird. Nature Communications. 2013-10-08, roá. 4, áûÙs. 1, s. 2554. Dostupnûˋ online [cit. 2021-12-28]. ISSN2041-1723. DOI10.1038/ncomms3554. (anglicky)
ãUNDERHILL, L. G.; PRYS-JONES, R. P.; DOWSETT, R. J. The biannual primary moult of Willow Warblers Phylloscopus trochilus in Europe and Africa. Ibis. 1992, roá. 134, áûÙs. 3, s. 286ã297. Dostupnûˋ online [cit. 2021-12-27]. ISSN1474-919X. DOI10.1111/j.1474-919X.1992.tb03811.x. (anglicky)
ãHEDENSTRûM, Anders. Adaptations to migration in birds: behavioural strategies, morphology and scaling effects. Philosophical Transactions of the Royal Society B: Biological Sciences. 2008-01-27, roá. 363, áûÙs. 1490, s. 287ã299. Dostupnûˋ online [cit. 2021-12-27]. DOI10.1098/rstb.2007.2140. PMID17638691. (anglicky)
ã ab Miliardy ptûÀké₤ se vracejûÙ z teplû§ch krajin, nákteéûÙ péekonajûÙ i tisûÙce kilometré₤. áT24 [online]. [cit. 2022-01-27]. Dostupnûˋ online.
ãHAHN, Steffen; BAUER, Silke; LIECHTI, Felix. The Natural Link between Europe and Africa: 2.1 Billion Birds on Migration. Oikos. 2009, roá. 118, áûÙs. 4, s. 624ã626. Dostupnûˋ online [cit. 2022-01-27]. ISSN0030-1299. (anglicky)
ã Cyprus poachers kill 2.3 million songbirds in 2016. phys.org [online]. [cit. 2022-01-27]. Dostupnûˋ online. (anglicky)
ã Ve StéedomoéûÙ pytlûÀci pozabûÙjûÙ roáná 25 milioné₤ ptûÀké₤. áeéÀtûÙ experti se snaéƒûÙ situaci zvrûÀtit. ct24.ceskatelevize.cz [online]. áT24 [cit. 2022-01-28]. Dostupnûˋ online.
ãLEWIS, David. We the Navigators: The Ancient Art of Lanfinding in the Pacific. Honolulu, Hawaii: Universitz of Hawaii Press, 1994. Dostupnûˋ online. ISBN9780824158231. S. 214ã216. (anglicky)
ã abcCROWE, Andrew. Pathway of the birds: the voyaging achievements of Máori and their Polynesian ancestors. Auckland, New Zealand: David Bateman LTD, 2019. ISBN9781869539610. S. 142ã152. (anglicky)
ãRICHTER-GRAVIER, Raphael. Manu narratives of Polynesia A comparative study of birds in 300 traditional Polynesian stories. Dunedin, New Zealand, 2019 [cit. 2021]. PhD Thesis. University of Otago. VedoucûÙ prûÀce Michael Reilly. Dostupnûˋ online.
ãSZABO, M. J. Pacific golden plover [online]. New Zealand Birds Online, 2013 [cit. 2021-12-09]. Dostupnûˋ online. (anglicky)
ã abCanoe Navigation, Locating land [online]. Te Ara: The Encyclopedia of New Zealand [cit. 2021-12-09]. Dostupnûˋ online. (anglicky)
ã abFONE, Martin. Curious Questions: How did a stork with a spear through its neck solve the mystery of the migration of birds? [online]. Country Life, 2020-01-11 [cit. 2021-12-10]. Dostupnûˋ online. (anglicky)
ãReview: The Wisdom of Birds by Tim Birkhead [online]. 2008-10-17 [cit. 2021-12-10]. Dostupnûˋ online. (anglicky)
ãHARRISON, Thomas P. Birds in the Moon. S. 323ã330. Isis [online]. 1954-12 [cit. 2021-12-10]. Roá. 45, áûÙs. 4, s. 323ã330. DOI10.1086/348354. (anglicky)
ãBEWICK, Thomas. History of British Birds. Newcastle: Beilby and Bewick, 1847. S. 300ã303. (anglicky)
ãBROSNAN, Elise. Bird of the Month: The Pfeilstorch [online]. Audubon Society of Northern Virginia, 2019-09-01 [cit. 2021-12-10]. Dostupnûˋ online. (anglicky)
ã abDer Pfeilstorch in der Zoologischen Sammlung [online]. University of Rostock, 2003 [cit. 2021-12-10]. S. 9ã10. (námecky)
ãInstructions for observing nocturnal bird migration by moon-watching [online]. University of Arizona, 2004 [cit. 2021-12-29]. Dostupnûˋ v archivu poéûÙzenûˋm dne 2021-12-29. (anglicky)
ãPREUSS, Niels Otto. Hans Christian Cornelius Mortensen: Aspects of his life and of the history of bird ringing [online]. Duglebjerg, Denmark: 2000 [cit. 2021-12-11]. Dostupnûˋ online. (anglicky)
ã abOVáIARIKOVû, S.. Metody sledovûÀnûÙ migrace zvûÙéat se zamáéenûÙm na telemetrii. 2020. FAKULTA VETERINûRNû HYGIENY A EKOLOGIE: ûstav biologie a chorob volná éƒijûÙcûÙch zvûÙéat. Dostupnûˋ online.
ã abKLVAéA, Petr; CEPûK, Jaroslav. Kudy a kam ptûÀci lûˋtajûÙ. Vesmir.cz [online]. 2018 [cit. 2021-12-29]. Dostupnûˋ online.
ãFrequently Asked Questions about GPS Transmitter (NC99) [online]. The Wildlife Center of Virginia [cit. 2022-01-28]. Dostupnûˋ v archivu poéûÙzenûˋm dne 2022-01-28. (anglicky)
ãBREZINA, Ivan. MigrujûÙcûÙ ptûÀci chytée vyuéƒûÙvajûÙ kaéƒdou péûÙleéƒitost k péeéƒitûÙ. I kosi. iDNES.cz [online]. 2019-03-22 [cit. 2021-12-29]. Dostupnûˋ online.
ã abHALLWORTH, Michael T.; MARRA, Peter P. Miniaturized GPS Tags Identify Non-breeding Territories of a Small Breeding Migratory Songbird. Scientific Reports. 2015-06-09, roá. 5, áûÙs. 1, s. 11069. Dostupnûˋ online [cit. 2022-01-27]. ISSN2045-2322. DOI10.1038/srep11069. PMID26057892. (anglicky)
ãNILSSON, Cecilia; DOKTER, Adriaan M.; SCHMID, Baptiste; SCACCO, Martina; VERLINDEN, Liesbeth; BûCKMAN, Johan; HAASE, Gû¥nther. Field validation of radar systems for monitoring bird migration. S. 2552ã2564. Journal of Applied Ecology [online]. 2018 [cit. 2021-12-29]. Roá. 55, áûÙs. 6, s. 2552ã2564. DOI10.1111/1365-2664.13174. (anglicky)
ãJOHNSON, Michael. International Space Station Images Trace Bird Migrations [online]. NASA, 2021-03-01 [cit. 2021-12-29]. Dostupnûˋ v archivu poéûÙzenûˋm dne 2021-12-08. (anglicky)
ãEMLEN, Stephen T.; EMLEN, John T. A Technique for Recording Migratory Orientation of Captive Birds. The Auk. 1966-07-01, roá. 83, áûÙs. 3, s. 361ã367. Dostupnûˋ online [cit. 2021-12-29]. ISSN1938-4254. DOI10.2307/4083048. (anglicky)
ãPROCHûZKA, Petr. Analû§za stabilnûÙch izotopé₤ ã alternativnûÙ metoda studia migrace ptûÀké₤. S. 3ã21. Sylvia [online]. áSO, 2006. Roá. 42, s. 3ã21. Dostupnûˋ online.
ãWorld Migratory Bird Day [online]. [cit. 2021-12-30]. Dostupnûˋ online. (anglicky)
ãHORTON, Kyle G.; LA SORTE, Frank A.; SHELDON, Daniel. Phenology of nocturnal avian migration has shifted at the continental scale. Nature Climate Change. 2020-01, roá. 10, áûÙs. 1, s. 63ã68. Dostupnûˋ online [cit. 2021-12-28]. ISSN1758-6798. DOI10.1038/s41558-019-0648-9. (anglicky)
ã abJENNI, Lukas; KûRY, Marc. Timing of autumn bird migration under climate change: advances in longãdistance migrants, delays in shortãdistance migrants. Proceedings of the Royal Society of London. Series B: Biological Sciences. 2003-07-22, roá. 270, áûÙs. 1523, s. 1467ã1471. Dostupnûˋ online [cit. 2021-12-28]. ISSN0962-8452. DOI10.1098/rspb.2003.2394. PMID12965011. (anglicky)
ãORELLANA-MACûAS, Josûˋ; BAUTISTA, Luis; MERCHûN, Daniel. Shifts in crane migration phenology associated with climate change in southwestern Europe. Avian Conservation and Ecology. 2020-05-08, roá. 15, áûÙs. 1. Dostupnûˋ online [cit. 2021-12-28]. ISSN1712-6568. DOI10.5751/ACE-01565-150116. (anglicky)
ã abcClimate change may be why birds are migrating earlier across the United States [online]. 2019-12-17 [cit. 2022-01-18]. Dostupnûˋ online. (anglicky)
ãDUNN, Peter O; WINKLER, David W. Climate change has affected the breeding date of tree swallows throughout North America. Proceedings of the Royal Society of London. Series B: Biological Sciences. 1999-12-22, roá. 266, áûÙs. 1437, s. 2487ã2490. Dostupnûˋ online [cit. 2022-01-18]. DOI10.1098/rspb.1999.0950. PMID10693819. (anglicky)
ãBOTH, Christiaan; BOUWHUIS, Sandra; LESSELLS, C. M. Climate change and population declines in a long-distance migratory bird. Nature. 2006-05, roá. 441, áûÙs. 7089, s. 81ã83. Dostupnûˋ online [cit. 2021-12-28]. ISSN0028-0836. DOI10.1038/nature04539. (anglicky)
ãLAWRENCE, Kieran B.; BARLOW, Clive R.; BENSUSAN, Keith. Phenological trends in the pre- and post-breeding migration of long-distance migratory birds. Global Change Biology. 2022, roá. 28, áûÙs. 2, s. 375ã389. Dostupnûˋ online [cit. 2022-01-27]. ISSN1365-2486. DOI10.1111/gcb.15916. (anglicky)
ãVISSER, Marcel E.; PERDECK, Albert C.; BALEN, JOHAN H. Van. Climate change leads to decreasing bird migration distances. Global Change Biology. 2009, roá. 15, áûÙs. 8, s. 1859ã1865. Dostupnûˋ online [cit. 2022-01-18]. ISSN1365-2486. DOI10.1111/j.1365-2486.2009.01865.x. (anglicky)
ãZIMOVA, Marketa; WILLARD, David E.; WINGER, Benjamin M. Widespread shifts in bird migration phenology are decoupled from parallel shifts in morphology. Journal of Animal Ecology. 2021, roá. 90, áûÙs. 10, s. 2348ã2361. Dostupnûˋ online [cit. 2021-12-28]. ISSN1365-2656. DOI10.1111/1365-2656.13543. (anglicky)
ãPANUCCIO, Michele; MARTûN, Beatriz; MORGANTI, Michelangelo; ONRUBIA, Alejandro; FERRER, Miguel. Long-term changes in autumn migration dates at the Strait of Gibraltar reflect population trends of soaring birds. S. 55ã65. Ibis [online]. 2017-01 [cit. 2021-12-15]. Roá. 159, áûÙs. 1, s. 55ã65. DOI10.1111/ibi.12420. (anglicky)
ãLA SORTE, Frank A.; HOCHACHKA, Wesley M.; FARNSWORTH, Andrew; DHONDT, Andrûˋ A.; SHELDON, Daniel. The implications of midãlatitude climate extremes for North American migratory bird populations. Ecosphere [online]. 2016-03. Roá. 7, áûÙs. 3. DOI10.1002/ecs2.1261. (anglicky)
ã abMigratory Birds in Culture [online]. World Migratory Birds Day (UNEP/CMS) [cit. 2022-02-19]. Dostupnûˋ online. (anglicky)
ã abSAX, Boria. Bird Migration and Human Culture [online]. [cit. 2022-02-19]. Dostupnûˋ online. (anglicky)
BERTHOLD, P., 2001. Bird migration: a general survey. Oxford: Oxford University Press. Dostupnûˋ online. ISBN9780198507871. (anglicky)
LINCOLN, Frederick C., 1935. Migration of Birds: Circular 16. Manhattan, Kansas, US: U.S. Fish and Wildlife Service. Dostupnûˋ online. (anglicky)
NEWTON, Ian, 2008. The migration ecology of birds. Amsterdam: Elsevier-Academic Press. ISBN978-0-12-517367-4. (anglicky)
RAPPOLE, John H., 2013. The avian migrant: the biology of bird migration. New York: Columbia University Press. Dostupnûˋ online. ISBN978-0-231-14678-4. (anglicky)
Film
Nedej se: [blok obáanskûˋ a investigativnûÙ éƒurnalistiky o éƒivotnûÙm prostéedûÙ]. Cesty taéƒnû§ch ptûÀké₤. Dokument áR. 2022. Reéƒie Pavel Jurda. Premiûˋra 16. éûÙjna 2022 11.55ã12.20 áeskûÀ televize (áT2).



















.jpg)


_(14763687525).jpg)








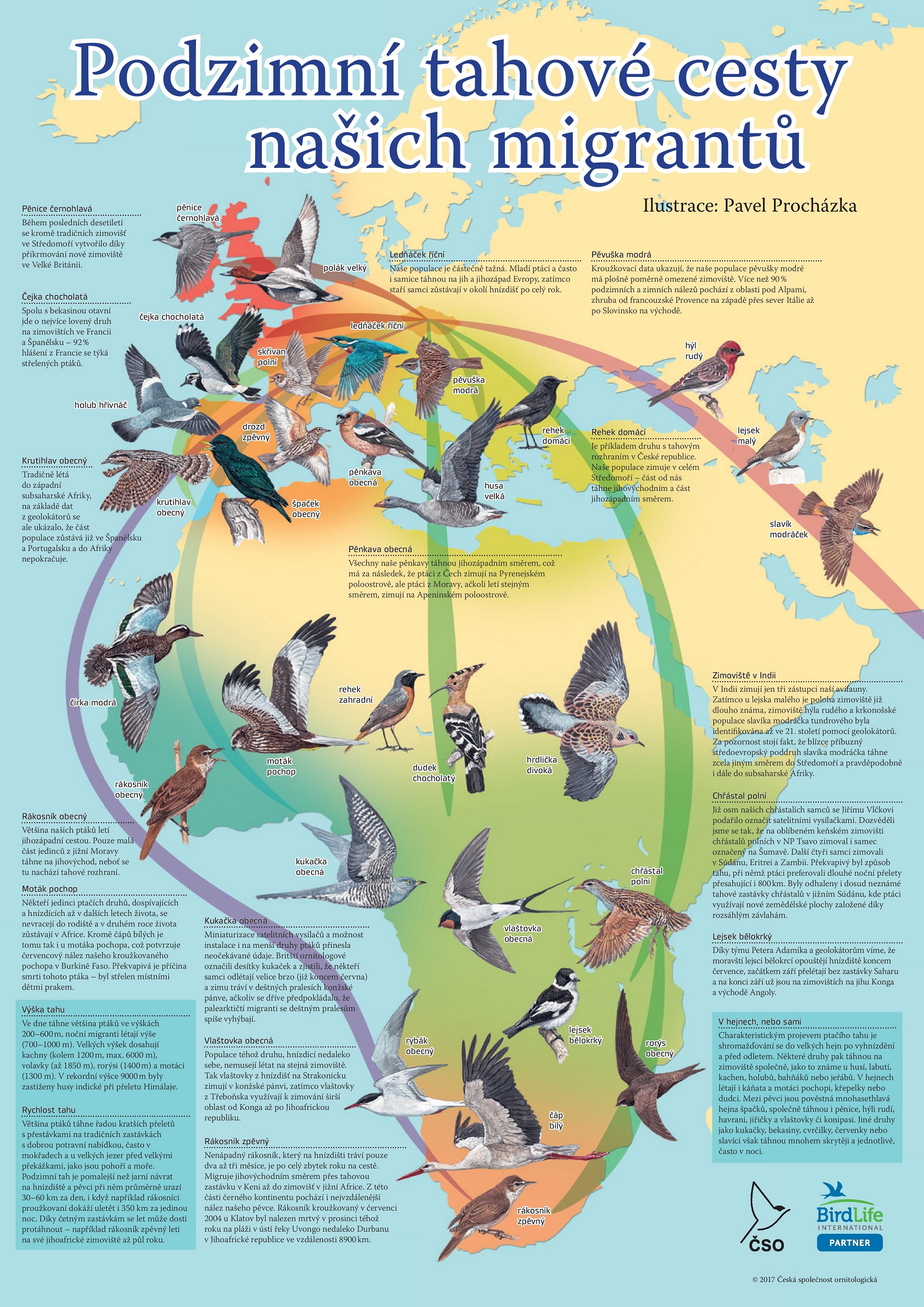{kind=link}
