ŠÊšÇښNJŠÀššš¡ÚÊ š¡š¯ÚÚ´šõ¯ õ¯šŠÑÚÇŠšÏ šŠ ATP š š˜šýÇš Š°çÚˋšýÇŠË¥ šÇŠÈ˜ Šˆ´šç. PDB ID:1UN9[ 1] ÚÊŠÊšÇšÊ (ššÇ : kinase )Š ATP š õ¯š õ° šŠšÏ š¡š¯ õý¯Úˋ ŠÑšš š¡š¯õ¡¯ ŠË¥ Ú¿š õ¡¯šÏ š š Š˜ÚŠ š¡š¯Ú ŠË¥ šÇŠÏÊ ÚŠ Ú´š šÇŠÊ. š¡š¯ÚÚ´š (ÓÕ
¡ÍÕ
çÓÇ ), ÚÊŠšš Š¥õ° Š ÚŠÊ. Úʊʚǚʊ š¡š¯õ¡¯š šÇÚ´š ŠÑŠËš š¥šÂ
šÇŠÊ. Úʊʚǚʊ Š˜Çõ¡¯ š¡š¯õ¡¯ŠË¥ ššˋšýÇš ŠÑõ¯ÚŠ Š¯šš ŠÏÊõ¯ÚŠ õ¯š¡š¯ŠÑÚÇÚ´š Š, š¡š¯õ¡¯ŠË¥ š õݯڊ Š¯šš ŠÏÊõ¯ÚŠ š¡š¯õ¯šŠÑÚÇÚ´š š õ瘊ÑÚš˜š¥ ÚŠÊ. Š´Š¯ÝšÏ , šÏšÏ ŠŠ ÚšÚŠ˜¥ š š¡š¯Ú šÚš Š¯Š¥ ÚšÝ, Š¯ššÝ, ŠÊŠË¡ ŠÑšš õý¯ÚˋÚ š šŠ ŠËŠ Ë ŠÝšÇ Š°ÚÚŠÊ. Š¯Š¥š Úʊʚǚʊ Š˜¥šÏŠš˜ , š¡Ú˜ š Ú¡š Š˜ , ŠýšÙ Ú Š°Ú , š¡Ú˜ ššÀ , ŠÑŠ¿ Š¯ õñ¡ Š¯š šŠÏš š¡Ú˜ Š¯š õý§ŠÀšš ŠÏÊš¯ šÊšÚ šÙÚ š ÚŠÊ.
1954Š
šÏ š¥ŠÊŠ(Gene Kennedy)õ¯ š¿Çš š¡ (casein )š š¡š¯ÚÚŠ õ¯ Ú´šŠË¥ Š¯õý˜ÚšŠÊ. šÇ Ú´šõ¯ ATPŠË¥ šÇšˋÚš˜ ŠÊŠË¡ Š´Š¯ÝšÏš š¡š¯Ú ÚŠ Š¯šš šÇŠÏÊÚŠ Ú´š ŠÀš šçšÇŠÀ õ¡¯ŠÀŠ õýšÇŠÊ. 1956Š
š šŠŠ´¥Š Ú¥š
(Edmond H. Fischer)š šŠš Ú˜Š Š¡šÊ (Edwin G. Krebs)Š õ¯š¡š¯ŠÑÚÇÚ´š aš õ¯š¡š¯ŠÑÚÇÚ´š b š˜šÇš šÚ¡ Š°ÚšÇ š¡š¯Úš Úš¡š¯Úš šÚÇ ŠÏÊõ¯ŠŠÊŠ õýš Š¯õý˜ÚšŠÊ.[ 2] õ¯š¡š¯ŠÑÚÇÚ´š š¡š¯ÚÚ´š ŠÀ Šˆ
Šˆ
ŠšŠÊ. Šˆ Š
Úš š¡š¯ÚÚ´šŠË¥ ÚçÚ š Ú¡š Š˜õý§ŠÀš šý¨ ššŠÀš Š´Š¯ÝšÏ š¡š¯ÚÚ´š A(PKA)õ¯ õ¯š¡š¯ŠÑÚÇÚ´š š¡š¯ÚÚ´šŠË¥ š¡š¯ÚÚŠÊŠ š˜šÊšÇ Š¯ÚšÀŠÊ. ŠÚ PKAõ¯ õ¡ŠÎ˜š§õý ÚˋšÝÚ´š (glycogen synthase )ŠË¥ š ÚÇÚŠÊŠ õýšÇ šŠ ÊšÀš¥Šˋ¯, šÇŠ š¡š¯ÚŠË¥ ÚçÚš˜ š ÚÇ ššˋš ÚŠ šçšÇš šššÇŠÊ. 1969Š
š Š šÊÚ¯ ŠÎ˜Š (Lester Reed)Š Ú¥ŠÈ´Š¡š¯ ÚššÚ´š õ¯ š¡š¯ÚŠšÇ ŠÑÚšÝÚŠŠÊŠ õýš Š¯ÚŠÇšÇ, š¡š¯Úõ¯ õ¡ŠÎ˜š§õý Šš˜Š¢ŠÏ šŠŠ¥ ŠÊŠË¡ Šš˜ õý§ŠÀšŠ šÀ¯š ššˋš ÚŠÊŠ Š´šŠË¥ š õ°çÚšŠÊ.[ 3] Ú¯ ŠÙõÝÇ (Tom Langan)š PKAõ¯ ÚšÊÚÊ H1š š¡š¯ÚÚŠˋ¯, Ú´šõ¯ šŠ Š´Š¯ÝšÏŠ š¡š¯ÚŠË¥ ÚçÚÇ šÀ¯š ŠŠÊõ° š õ¡¯ÚšŠÊ.[ 4] š¿¥Šˆ´ŠŠÎ¯ ššÀÇšÝ Š´Š¯ÝšÏ š¡š¯ÚÚ´š õ¯ Š¯õý˜Ššš¥Šˋ¯, Š´Š¯ÝšÏš ÚŠš šŠ₤¡Š
¡š¯ šõ¡¯ šÇšš¥ŠÀ š¡š¯ÚŠ š šššÇ šŠ ÊšÀŠÊ. 1990Š
Šš ŠÊšÇš MAPK/ERK õý§ŠÀ , JAK š¡š¯ÚÚ´š (Ú¯ŠÀš š¡š¯ÚÚ´ššÀÝ), õñ¡ŠÎ˜õ° PIP3 ššÀÇšÝ š¡š¯ÚÚ´š š Ú¡š Š˜õý§ŠÀ ŠÝšÇ õ°šÚÇš Š¯ÚšÀŠÊ.
š¡š¯ÚÚ´šŠ õ¡¯šÏš Š¯Š¥ Š´Š¯ÝšÏ š¡š¯ÚÚ´š, šÏšÏ š¡š¯ÚÚ´š, ÚšÚŠ˜¥ š¡š¯ÚÚ´š ŠÝ ŠÊšÚ õñ¡ŠÈ¿š¥ŠÀ ŠŠŠÊ. š¡š¯ÚÚ´šŠ Š¯Ú
ŠÎ˜ššš õ°¯ÚÀšÇ, õ°ÊšÑˋ, Ú˜š ŠËš šÇŠËÇõ¡¯õ¿šÏ ŠÊšÚ šÂ
šš Š¯õý˜ŠŠÊ.[ 5] [ 6] õ° ŠÎ˜Ú AMP š õý¯ÚˋÚš˜ šÀ¯š ŠŠÊ.[ 7]
š¡š¯ÚÚ´šŠ õ° šŠšÏŠÑš, šŠË¥ ŠÊšÇ ATP š õ¯š ŠÑšš š¡š¯ ššˋõ¡¯ ŠË¥ õ¡¯šÏ ŠÑšŠÀ šÛõ¡¯Š Š¯šš ŠÏÊõ¯ÚŠÊ. šÇ Š¯ššŠ õ° šŠšÏŠË¥ õ¯šÏŠ š¡š¯Š˜Çšõý¯ÚˋšÇ õÇš˜Úõ¡¯ ŠŠ˜¡š š¡š¯ÚÚ´šŠ š¡š¯Š˜Çšõý¯Úˋš šš ÚÚŠ šÙÚ š ÚŠÊ. š¡š¯ÚÚ´šŠ õ¡¯šÏõ°¥ š¡š¯õ¡¯ŠË¥ ÚšÝ šŠÎ˜ š š š Úõý Š¯¯šÇÚš˜ Š¯š šŠ ŠË¥ Šš¡ŠÊ. šš ÚŠË¥ ŠÊ š¡š¯õ¡¯š šÚ¡ššˋÚš˜ ŠÚŠŠ š šÇšÚŠ ŠŠÑŠÑš õý§š¯ šš ÚŠË¥ ŠÊ šŠ₤¡Š
¡š¯š ÚçÚÇ š š õ¡¯š š¥ŠÀ šš ÚŠŠÊ. ŠˆŠˆ š¡š¯ÚÚ´šŠ õ¡š Š°ÇšÀ¯š¡š ŠË¥ šÇšˋÚš˜ š¡š¯õ¡¯š Š¯¯šõý¯ÚˋÚŠÊ.
š¡š¯ÚÚ´šŠ š¡Ú˜ŠÇ š Ú¡š Š˜ (signal transduction )š õÇŠýšÚ šÚËš Š₤¡š¿õ° š¡Ú˜ ŠÇŠÑš Š°çšÀÚ õ¡¯šŠÊš šÀ¯š ÚŠÊ. šÇŠÊ ŠÑšŠ š¡š¯ÚŠË¥ ÚçÚš˜ ÚšÝšÇ õ¯ÚŠõݯŠ š ÚÇŠŠˋ¯, ŠÊŠË¡ ŠÑšš šÚ¡ššˋÚŠ ŠËŠ ËšÇ šÀ¯š ŠŠÊ. ŠÏš š¡š¯ÚÚ´šõ¯ Úõý§ šÀ¯õÝǚNJ š Ú¡š Š¯Š¥ Š¯šÚõ¡¯ ŠŠ˜¡š š¡Ú˜Š šÚˋš Š¯Š¥ š¡Ú˜ ŠÇš ŠÑšŠË¥ Úçš Ú š šŠÊ. š¡š¯ÚÚ´šš õ¡¯ŠËš ššÊÚŠ Šš¯Š°šÇŠ õ¡¯ŠËš š£Š Šš¯Š°šÇõ¯ Š¯šÚŠˋÇ š [ 8] neuroblastoma ), šýššŠšÝ šÇŠšÊšÀ¯(spinocerebellar ataxia type 14 ), Š˜Çõ¯ŠÏõ¡ŠÀŠÑŠÎ¯ÚšÎ(agammaglobulinaemia ) ŠÝ[ 9]
š¡š¯ÚÚ´šõ¯ šÇŠÏÊ ššˋÚŠ š¥Š¯š š¡ Š¯š
š Ú¡š Š˜õý§ŠÀš õ¯š. š¯õÇŠ Š´Š¯ÝšÏ šÊ ŠÊšõ¯ š¡š¯ÚÚ´ššÇŠˋ¯ MAPKš JAK õ¯š Š´Š¯ÝšÏ š¡š¯ÚÚ´šš PI3K õ¯š šÏšÏ š¡š¯ÚÚ´šõ¯ Ú˜Ú´ŠšÇ šŠÊ. Š´Š¯ÝšÏ š¡š¯ÚÚ´š (Protein kinase )Š Š´Š¯ÝšÏš š¡ŠÎ¯, Ú¡Š šÊŠ, ښNJÀš , ÚšÊÚ¯Š šõ¡¯ŠË¥ š¡š¯ÚÚŠÊ. š¡š¯ÚŠË¥ ÚçÚš˜ Š´Š¯ÝšÏš õ¡¯ŠËš ŠÊŠ¯ˋŠˋÇš¥ŠÀ Š°ÚÚŠÊ. Š´Š¯ÝšÏš ښݚ šÎõ¯šÚÊõݯŠ õ¯ššÚÊõ° , šš ÚÚõݯŠ ŠÑÚÇŠË¥ šÚ ÚššÇ Šõ¡¯Š ÚŠˋ¯, Ú¿š Ú š¡Ú˜ õç˜Úš šš¿šÚÊõ¡¯Š Úõ° , ŠÊŠË¡ Š´Š¯ÝšÏõ°¥š šÚ¡ššˋš õ¯šÚõݯŠ õçŠÚ š šŠÊ. Š´Š¯ÝšÏ š¡š¯ÚÚ´šŠ š¡š¯ÚÚ´šš ŠŠÑŠÑš š¯´šÏÚŠˋ¯ ŠŠÎ˜ š¯õ瘚 ŠššÇ ŠŠÊ.[ 6]
š¡Ú˜ Š´Š¯ÝšÏš šŠÏš õ°çš õý¯Úˋš ŠššÇšÏŠÏ š¡š¯ÚŠ¯ššýŠ¥ õ¯šÙš š¡ õ°çš õý¯Úˋš ŠÏšÏ šŠÊ. šÇŠÀ š¡Úš˜ Š´Š¯ÝšÏš š¡š¯Úõ¯ šÀ¯š š õ¡¯ŠËš õ¯Š õýšÇŠÊ. ŠÊŠË¡ šŠÎ˜ š
šýÇšÝ šÀ¯š šÇš¡šŠ Š´Š¯ÝšÏõ°¥ õ°çš õý¯ÚˋÚ š šŠ Š¯ˋŠýšÇ ŠÏš¥Š₤ŠÀ Š´Š¯ÝšÏ õ¡¯ŠËš šÀ¯š Ú š šŠ õ¯ŠËšÝš Š˜ÇõÑŠ˜ÇšÏÚŠÊ. šŠš Ú˜Š Š¡šÊŠ ÚÚ´šÊ Úõ° õ¯š¯šš ŠÊŠË¡ šŠÎ˜ š
šýÇšÝ šÀ¯š š š¡Ú˜ ŠÇŠÑšš š¥šÇŠŠ š Ú¡š Š¯šÚõ¡¯ šÚš˜ š¥šÇŠŠ Š¯ŠˋÇ, š¡š¯ÚŠ š¡Ú˜ š¡ŠÑŠÀŠÑÚ¯š š Ú¡š ŠÚÇ š¥šÇŠŠŠÀ šÏÚÚšŠÊõ° šÈ¥šËÚšŠÊ. šÚ皊˜¥ š Š¿Úš˜ šÏÚ皊˜¥ š ÚÙŠš š Ú¡š Š¯šÚŠŠÀ šÏÚÚ Š°çšÀÚ š¡Ú˜šÇõ¡¯ ŠŠ˜¡š, šÇ šÈ¥šËš Š´Š¯ÝšÏ š¡š¯Úõ¯ šÏÚ皊˜¥šš Š Š¿ŠýÚõý š¥šÇŠŠÊŠ õýõ°¥ š¥š¿ÚŠÊ.[ 7]
š˜šÇÚNJί ššÀÇšÝ š¡š¯ÚÚ´š (Cyclin-dependent Kinase ; CDKs)Š š¡Ú˜šÈ¥õ¡¯ šÀ¯š õ°¥ õÇŠ ´Š š¡š¯ÚÚ´šŠÊšÇŠÊ. ŠÊŠË¡ Š´Š¯ÝšÏš š¡ŠÎ¯ Ú¿š Ú¡Š šÊŠ šõ¡¯ŠË¥ š¡š¯ÚښϊÏ, Š¯Šš š˜šÇÚNJί Š´Š¯ÝšÏõ°¥ Š´¥š õý¯ÚˋÚš˜š¥ ښݚ Š°Çš¡ŠÊ.[ 10] CDK ÚšÝÚ š¡š¯ÚÚ´š )š š¡š¯ŠÑÚÇÚ´š(e.g. Cdc25 )š šÀ¯š š Š¯ŠŠÊ.[ 11] [ 12] [ 13] [ 13]
š š˜ŠÑšÇ ÚšÝÚ Š´Š¯ÝšÏ š¡š¯ÚÚ´š(Mitogen-Activated Protein Kinases ; MAP kinase; MAPKs)Š š¡ŠÎ¯/Ú¡Š šÊŠ š¡š¯ÚÚ´ššÀÝš¥ŠÀ ŠÊšÚ š¡Ú˜š¡ šÝšË š Ú¡š Š¯šÚŠÊ. šÝšË Ú¡ŠËÇŠˆ˜ šÇŠ šÚ¥šÝšËš¡š , ÚšÚ š Š šÝšËš¡š , š¡šŠÎ¯ š Šˆ´Š š š˜ŠÑšÇ šõñ¿š¥ŠÀ MAPK õý§ŠÀš š¯õ°ŠŠÊ. ššˋšýÇ ššÊšš MAPK õý§ŠÀõ¯ ÚšÝÚŠŠˋÇ š Ú¡š Š˜ š¯šŠ¯ššÇ õ¯šŠõ° Ras GTPase Š GDPŠË¥ GTPŠÀ õçÚÚŠÊ. šÇšÇš RasŠ Raf š¡š¯ÚÚ´š(MAPKKK)ŠË¥, Raf š¡š¯ÚÚ´šŠ š š˜ŠÑšÇ ÚšÝÚ Š´Š¯ÝšÏ š¡š¯ÚÚ´š š¡š¯ÚÚ´š(Mitogen-activated Protein Kinase Kinase ; MAPKK; MEK)ŠË¥, MEKŠ MAPK(ERK)ŠË¥ ÚšÝÚÚŠÊ. MAPKŠ šÇšÇš š š˜ š ŠýšÙ š šÀ¯š ÚŠÊ. Rafš MAPKõ¯ š¡ŠÎ¯/Ú¡Š šÊŠ š¡š¯ÚÚ´šš¡Š¯ Š¯Úš˜ MAPKKŠ ښNJÀš /Ú¡Š šÊŠ š¡š¯ÚÚ´ššÇŠÊ.
ŠÊšÚ š š˜ŠÑšÇ š Ú¡õ¯ MAPK š¯šŠ¯šš ÚçÚš˜ š¡Ú˜ šÝšËõ°¥ ŠÑÚŠË¥ šÇšÏÚŠÊ. MAPKŠ š š˜š¡šŠË¥ šÏôñõ¯š š š¥ŠÀ šÀ¯š ÚŠÊ. šÈ¥š š š˜ Úš š ATF-2, Chop, c-Jun, c-Myc, DPC4, Elk-1, Ets1, Max, MEF2C, NFAT4, Sap1a, STATs, Tal, p53, CREB, õñ¡ŠÎ˜õ° MycŠË¥ Ú˜Ú´ÚŠÊ. ŠÚ ŠÎ˜Š°Çš ŠŠ´ššýÇš S6 š¡š¯ÚÚ´šŠË¥ š¡š¯ÚÚš˜ ŠýšÙš šÀ¯š ÚŠÊ. MAPK š¯šŠ¯šš šÇõ¡¯ õý§ŠÀŠË¥ šÇŠÈ´Š Ras, Sos, šÚ¥šÝšËš¡š ššˋšýÇ(EGF receptor) ššýÇŠ š¡š¯Ú Ú š šŠÊ.[ 14]
MAPK õý§ŠÀŠ ššš š¥ŠÀ šÈ¥Šˆˋš Š¯õ° šŠÊ. MAPK õý§ŠÀõ¯ õÇš˜ÚŠ š¡Ú˜ õ¡¯šš š¡Ú˜šÝšËš Úçš Úõ¡¯ ŠŠ˜¡š MAPK õý§ŠÀš Šš¯Š°šÇõ¯ šõ¡¯ŠˋÇ š¡Ú˜ŠÑÚ, šÎš, ššÀÇ, š¡Ú˜ššÇ (apoptosis )š ŠÚ šÀ¯š ššˋšÇ Š°šÏŠõ° Š¯Š¥š šÂ
ššÇ ššÝŠ š šŠÊ.[ 14]
šÏšÏ š¡š¯ÚÚ´š (Lipid kinases )Š šÚšÏŠÏ Š¢ŠÏ šŠŠ¥ š¡Ú˜šõ¡¯õÇ ŠÏš šŠ šÏšÏš š¡š¯ÚÚŠÊ. š¡š¯õ¡¯š ŠÑõ¯ŠÀ šÏšÏš Š¯ššÝõ°¥ šš¿õ¯ Š˜Š¥šÏõ° š Ú¡ š Š˜š šÇšˋŠ š šŠÊ.
Ú˜šÊÚÚ¯ŠšÇŠ
¡šÚ´ š¡š¯ÚÚ´š(Phosphatidylinositol kinases )Š Ú˜šÊÚÚ¯ŠšÇŠ
¡šÚ´ (phosphatidylinositol )š š¡š¯ÚÚš˜ Ú˜šÊÚÚ¯ŠšÇŠ
¡šÚ´ 3,4-šÇš¡š¯(phosphatidylinositol 3,4-bisphosphate , PI(3,4)P2 ), Ú˜šÊÚÚ¯ŠšÇŠ
¡šÚ´ 3,4,5-š¥š¡š¯(phosphatidylinositol 3,4,5-trisphosphate , PIP3 ), õñ¡ŠÎ˜õ° Ú˜šÊÚÚ¯ŠšÇŠ
¡šÚ´ 3-š¡š¯(phosphatidylinositol 3-phosphate , PI3P) ŠÝš ŠÏŠ ŠÊ. Ú˜šÊÚÚ¯ŠšÇŠ
¡šÚ´ š¡š¯ÚÚ´šš šŠÀŠ Ú˜šÊÚ˜šÇŠ
¡šÚ¯Š 3-š¡š¯ÚÚ´š(phosphoinositide 3-kinase , PI3K), Ú˜šÊÚ˜ÚÚ¯ŠšÇŠ
¡šÚ´-4-š¡š¯ 3-š¡š¯ÚÚ´š(phosphatidylinositol-4-phosphate 3-kinase , õñ¡ŠÎ˜õ° Ú˜šÊÚÚ¯ŠšÇŠ
¡šÚ´-4,5-šÇš¡š¯ 3-š¡š¯ÚÚ´š(phosphatidylinositol-4,5-bisphosphate 3-kinase )õ¯ šŠÊ. Ú˜šÊÚÚ¯ŠšÇŠ
¡šÚ´š š¡š¯Ú šÚŠ š¡šŠÎ¯ š Ú¡š Š˜õý§ŠÀš õ¯š š¡Ú˜š Ú¡š Š˜š šÊšÚ šÙÚ š Úõ° , š¡Ú˜ŠÇšÇš
(endocytosis ), š¡Ú˜š¡š šÑ(exocytosis Š¯ õ¡¯Ú ššÀšŠ šÙÚ š ÚŠÊ.[ 15] [ 16] š¡šŠÎ¯ š ÚÙšÝ š š Š¯Ú š šŠÊ.[ 17]
š¡šŠÎ¯šÇ ššˋšýÇš õý¯ÚˋÚŠˋÇ PI3Kõ¯ ŠÏš õý¯ÚˋÚš˜ Ú˜šÊÚÚ¯ŠšÇŠ
¡šÚ´ šÏšÏš š¡š¯ÚÚ š šŠÊ š¡š¯ÚÚ´šŠ šÇŠ
¡šÚ´ šš¯Úõ¡¯ŠË¥ Š š¿ÚçšÝš Š õý Úš˜ Š¯š šŠŠË¥ šÎõ¯šÚÊŠŠ¯, šÂ
šÂ
šŠ õ¡¯šõ°¥ õ¯šÇ šŠ₤¡Š
¡š¯ šõ¡¯š õ°š˜š˜š š¥Š¯ š¥õ¡¯ŠÀ ššˋÚõý Úš˜ šš¯Úõ¡¯šš ššŠË¥ š õݯڊÊ.[ 18] [ 18]
Ú˜šÊÚÚ¯ŠšÇŠ
¡šÚ´-3 š¡š¯ÚÚ´šš õ¡¯š. ATPš Ú˜šÊÚÚ¯ŠšÇŠ
¡šÚ´šÇ š¥Š¯ š¥õ¡¯ Bš Ššš Š¯š Š¯šÚš˜ Ú˜šÊÚÚ¯ŠšÇŠ
¡šÚ´-3-š¡š¯õ°¥ ADPŠË¥ šš¯ÚŠÊ.[ 18]
šÊÚõ° š š¡š¯ÚÚ´š(Sphingosine kinase )š šÊÚõ° š š šÊÚõ° š -1-š¡š¯(sphingosine-1-phosphate , S1P)š¥ŠÀ š ÚÚŠ Ú´ššÇŠÊ. šÊÚõ° šÏšÏš ŠÏš ŠŠÎ˜ šÀÇš˜ÚŠ ŠÏšÏšÏšÇŠÊ. šÊÚõ° š š¡š¯ÚÚ´šõ¯ ÚšÝÚŠŠˋÇ š¡Ú˜šÏšš šÚšÏŠÏš¥ŠÀ šÇŠÚš˜ ATPŠ GTPš ö° š¡š¯õ¡¯(š š¥ Š Ú¿š ŠÏšÏŠÏ š¡š¯õ¡¯)ŠË¥ šÊÚõ° š š¥ŠÀ š Š˜ÚŠÊ. S1P ššˋšýÇŠ GPCR ŠÀ S1PŠ G Š´Š¯ÝšÏ š Ú¡š Š˜š šÀ¯š Ú š šŠ ŠËŠ Ëš šÏŠŠÊ. õý¯õ°¥š š¥ŠÀ ERK, Rho GTPase, Rac GTPase š¡šÏšÏŠÑÚÇÚ´š C(phospholipase C , PLC) õñ¡ŠÎ˜õ° AKT/P13Kš õ¯š š¡Ú˜ŠÇ š¡šŠË¥ ÚšÝÚÚŠÊ. ŠÚ š¡Ú˜ ŠÇš Úš ŠÑšš Ú´õ°¥ŠË¥ Š¯ÚÚŠÊ. S1PŠ ÚšÊÚÊ Úšš¡Ú¡Ú´š (histone deacetylase , HDAC)š Úšš¡Ú¡ ښݚ šÏš š š¥ŠÀ š ÚÇÚŠÊ. Š¯ŠŠÀ Úš¡š¯ÚŠ šÊÚõ° š š š¡Ú˜ššÇš šÇšÏÚõ° Š¯Š¥š šÊÚõ° š š¡š¯ÚÚ´šš šÀ¯š š šÇÚÇÚŠ õýšÇ šÊšÚŠÊ. šÊÚõ° š š¡š¯ÚÚ´šõ¯ š¡Ú˜šÎšš šÇšÏÚõ° SK1(šÊÚõ° š š¡š¯ÚÚ´šš Ú šÂ
ŠË)šÇ Ú¿š š Úš ššš õ° ŠŠŠÀ šÀÇš˜ÚŠÊŠ š š Š₤¡ŠÈ´šÇ Š°Çš šÊÚõ° š š¡š¯ÚÚ´šõ¯ šš¡Ú˜š šÝšËš šÏššÚ˜ õ¯ŠËšÝšÇ šŠÊ. Ú˜š ŠŠ˜¥ š¡Ú˜šŠ SK1š SK2 Š šÂ
ŠËš šÊÚõ° š š¡š¯ÚÚ´šõ¯ šÀÇš˜ÚŠÊ. SK1šÇ SK2š Š¿Úš˜ Š Ú¿šÇš šÇõ° , õñ¡ŠÊš Š¯Ú š ÚŠ ššÇÚŠÊ. SK1š Ú, Š¿šË, ŠÎ¥Úõ瘚 Š¯ÚŠŠ Š¯ŠˋÇ SK2š š šËõ°¥ õ¯š¡Ú˜šš Š¯õý˜ŠŠÊ. šÇ Š š¡š¯ÚÚ´šŠ š¡Ú˜ššÀÇ, šÎš, ŠÑÚ Š¯ š¥šÎ õ°¥ š¯õÇšÇ ššÇ ÚÚšŠý š ŠššÇ ŠŠÊ.[ 19]
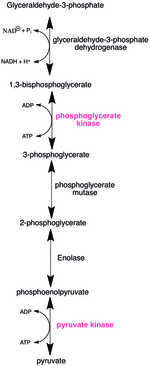
ÚšÚŠ˜¥š Š¿ Š´ŠÑšŠÀ ŠÑÚÇÚŠ šý¨ Š´õ°š¡ ÚÇŠ¿ õ°¥š ššŠ š¡š¯Úõ¯ Š Šý š¥šÇŠŠÊ ŠÏš Ú˜š ŠŠ˜¥šõý ÚšÚŠ˜¥š š¥š¥ šÇŠ ÚšŠš ŠŠÑŠÑš õ°çõ¡ÚŠÊ. š˜ŠÎ˜õ° Š¿ šš šŠšÏŠË¥ š£õ¡¯ šÚÇšŠ š¯š Š´Š¿ŠË ŠÀ ŠÑÚÇŠšÇ š¡Ú˜Šš˜ŠÀ ŠÊšÇõ¯š¥ ÚŠÊ. š¡š¯ÚÚ´šŠ õݯš Šˆ´Š Šš˜ õý§ŠÀšš šÊšÚ šÙÚ š ÚŠÊ. õñ¡ŠÎ¥š ÚÇŠ¿ (glycolysis )š Š ŠýšÏ¡ šõ¡¯ŠË¥ Š°Çš˜šÊŠÊ. 1,3-šÇš¡š¯ õ¡ŠÎ˜š¡ŠËÇš¯š Š˜Çšõý¯Úˋš ŠÑšš Úõ° Šš šŠšÏŠË¥ õ¯šÏŠÊ. 1,3-šÇš¡š¯ õ¡ŠÎ˜š¡ŠËÇš¯ š¡š¯ÚÚ´šŠ ADPŠË¥ šÇšˋÚš˜ 3-š¡š¯ õ¡ŠÎ˜š¡ŠËÇš¯õ°¥ ATPŠË¥ šš¯ÚŠÊ. ÚÇŠ¿š ŠÏšÏŠÏ õ°¥š šš Ú¥ŠÈ´Š¡š¯ š¡š¯ÚÚ´šŠ Ú˜šÊÚ˜šš˜Ú¥ŠÈ´Š¡š¯š š¡š¯õ¡¯ŠË¥ ADPŠÀ š Š˜Úš˜ ATPš Ú¥ŠÈ´Š¡š¯š šš¯ÚŠÊ.
ÚËšÚÊŠšš (ÚËšš¡š¯ÚÚ´š, Hexokinase )Š Ú˜ŠŠ¿šÇ šýš š¡Ú˜ ŠÇŠÀ ŠÊšÇõ¯õý ÚŠ š¥Š¯š š¡ Ú´ššÇŠÊ. ATPš õ¯ŠÏ š¡š¯õ¡¯ŠË¥ D-Ú˜ŠŠ¿š C6ŠÀ š Š˜Úš˜ Ú˜ŠŠ¿-6-š¡š¯š¥ŠÀ š ÚÚŠÊ. š¡š¯õ¡¯š šš ÚŠÀ š¡ÚÇ Ú˜ŠŠ¿-6-š¡š¯š š¡Ú˜šÏ ŠÇš Š´¡Š˜ÇŠËÇõ° ÚÇŠ¿ õ°¥š š¥ŠÀ ŠÊšÇõ¯ š šŠÊ. Úš¡š¯ÚŠ ÚÚš Ú˜ŠŠ¿š ŠÊš š¡Ú˜ŠÏš õݯš° š¡Ú˜ Š¯õ¿Ëš¥ŠÀ š§õý š šÑŠŠÊ.[ 20] nonspherocytic hemolytic anemia )šÇ šõ¡¡ š šŠÊ.[ 21]
š¡š¯õ°¥Š¿ š¡š¯ÚÚ´š (Phosphofructokinase , PFK)Š õ°¥Š¿-6-š¡š¯š õ°¥Š¿-1,6-šÇš¡š¯š¥ŠÀ š ÚÚŠÊ. šÇ Š¯šš ÚÇŠ¿š šÊšÚ šÀ¯š Š¯ššÇŠÊ. ATPŠ H+ , õñ¡ŠÎ˜õ° šÚ¡ŠËÇš¯ š ŠŠõ¯ Šš¥ŠˋÇ š¡š¯õ°¥Š¿ š¡š¯ÚÚ´šŠË¥ š ÚÇÚŠÊ. šÚ¡ŠËÇš¯ ŠŠõ¯ Šš¥ŠˋÇ ÚÇŠ¿šÇ š š šŠŠÀ š¥šÇŠõ° šŠÊŠ õýš šŠ₤¡ÚŠÊ. šŠ¯Š
¡š š¥š¡š¯š ŠŠõ¯ Šš¥ŠˋÇ š¡š¯õ°¥Š¿ š¡š¯ÚÚ´šš ÚšÝšÇ ŠššÏŠÊ. š 7Ú Š¿šš šËŠ°(glycogen strage disease Ú¿š Tarui's disease )Š š¡š¯õ°¥Š¿ š¡š¯ÚÚ´š š š šš Šš¯Š°šÇõ¯ šõý´ ÚšÝšÇ ŠÛššÀõ¡¯ ŠŠ˜¡š š Š¯ŠŠÊ.[ 22]
ŠÎ˜Š°ÇÚŠ¥Š¿ š¡š¯ÚÚ´šš ښݚŠÎ˜š õý¯ÚˋÚ, Ú´šš šš¯Š˜¥š¡ FMN(š¥šˆ§)õ°¥ ADP(šÊŠË¡šˆ§). PDB ID: 1N07.[ 23] š¡š¯ÚÚ´šŠ Š´Š¯ÝšÏ, šÏšÏ, ÚšÚŠ˜¥ šÇš¡š ŠÑššŠ ššˋÚŠÊ. ŠÇÚÇŠ šÊÚ¯Š šÚ¡š Úš õÇŠ ´Š Ú´šŠÀ ŠÇÚÇŠ šÊšŠ š¡š¯ š¡š¯ÚÚ´šš ŠÇÚÇŠ šÊšŠ šÇš¡š¯ š¡š¯ÚÚ´šõ¯ šŠÊ.[ 24] Ú˜Š šÚÇ , š¡š¯õ¡ŠÎ˜š¡ŠËÇš¯ , ŠÎ˜Š°ÇÚŠ¥Š¿ , ŠÊšÇښNJŠÀššš¡ÚÊ , šÚÇš¯ (shikimate ) ŠÝŠÝšÇ šŠÊ.
ŠÎ˜Š°ÇÚŠ¥Š¿ š¡š¯ÚÚ´š(Riboflavin kinase )Š ŠÎ˜Š°ÇÚŠ¥Š¿š š¡š¯ÚÚš˜ ÚŠ¥Š¿ Šˆ´Š
¡ŠÇÚÇŠ šÊÚ¯Š(flavin mononucleotide , FMN)š ŠÏŠ ŠÊ. šÇ Š¯šššŠ ŠÎ˜Š°ÇÚŠ¥Š¿šÇ ATP ŠÑšš õý¯ÚˋÚõ¡¯ š š š¡š¯ÚÚ´šš Š´¥š õý¯ÚˋÚš˜š¥ ÚŠÊ.[ 25] [ 25]
ŠÎ˜Š°ÇÚŠ¥Š¿ š¡š¯ÚÚ´šš õ¡¯š ŠÎ˜Š°ÇÚŠ¥Š¿ š¡š¯ÚÚ´šŠ š¡Ú˜šš šÊšÚ šÙÚ š ÚŠÊ. FMNš šÊšÚ Š°ÇšÀ¯š¡š šÇŠˋ¯, š¯ÚÚšŠ¯šš Š°ÇšÀ¯š¡šš¡ ÚŠ¥Š¿ šŠ¯Š ŠŠÇÚÇŠ šÊÚ¯Š(flavin adenine dinucleotide , FAD)š š õ瘚ýÇšÇõ¡¯Š ÚŠÊ. ŠˆŠˆ Ú´šŠ ŠÎ˜Š°ÇÚŠ¥Š¿š FMNš¥ŠÀ, FMNš FADŠÀ š¡š¯ÚÚŠ Š¯š Šˆ´ŠŠË¥ šÚÚ š šŠÊ.[ 26] [ 27] [ 28]
Ú¯Š₤¡Š š¡š¯ÚÚ´š(Thymidine kinase )Š ŠÇÚÇŠ šÊšŠ š¡š¯ÚÚ´š šÊ ÚŠŠÀ ŠÇÚÇŠ šÊšŠ š¡š¯ÚŠË¥ ŠÇŠ¿ÚŠÊ. šŠšš Š°ÇšÇŠ õýõ°¥ õ¯šÇ ATP ŠÑššš š¡š¯õ¡¯ŠË¥ Ú¯Š₤¡Šš š Š˜Úš˜ Ú¯Š₤¡Š š¥š¡š¯(thymidine monophosphate , dTMP)š šš¯ÚŠÊ. Ú¯Š₤¡Š š¡š¯ÚÚ´šŠË¥ Š¿ŠÀ₤Ú ŠÊŠË¡ ŠÇÚÇŠ šÊšŠš ŠÇÚÇŠ šÊÚ¯Š š¡š¯ÚÚ´šŠ ŠÇÚÇŠ šÊÚ¯Šš ššÊš šÀ¯š ÚŠ šÙÚ š ÚŠÊ.
Ú¯Š₤¡Š š¡š¯ÚÚ´šõ¯ šÇŠÏÊ ššˋ ÚŠ š šýÇ Š¯š dTMP ŠÑšŠË¥ ššÝÚ Úš Ú¯Š₤¡Šš¯ š¡š¯ÚÚ´šõ¯ dTMPš ššˋÚš˜ šÇš¡š¯ ÚÚš¡ dTDPŠË¥ ššÝÚõ° , Š ŠÊŠË¡ Ú´šõ¯ ššˋÚš˜ dTTPŠË¥ šš¯Úš˜ DNA ÚˋšÝ š šÇšˋŠŠÊ. šÎ Ú¯Š₤¡Š š¡š¯ÚÚ´šš ښݚ š¡Ú˜šÈ¥õ¡¯š Š¯š Ú š¯õÇšÇ šš¥Šˋ¯ šš¡Ú˜šš ÚšÝšÇ šÎõ¯Ú š šõ° ,[ 29] [ 30] Š₤¡Ú š§ŠŠÎ˜š DNA õý¯Ú šÎÚõ篚 õýˆš š šŠÊ.[ 31]
õ¯Š§Ú ÚÊŠšš õý¯Ú(galactokinaseÓ¥¤ð¿)š õ¯Š§Ú ÚÊŠšš õ¯ š šýš š¥ŠÀ õý¯ÚŠ Šš˜ šËš šÇŠÊ. ÚšÀ šš õ¯Š§Ú šÊšÊ šš¿õ¯ šÎõ¯Šõ° Š¯ÝŠÇšË, õ¯ šÂ
Š, š š šÏšýÇ Š¯šš šÎšš Š°Çš¥ššŠÊ.
õ¡ŠÈ´š§ÚÊŠšš (glucokinase)Š õ¯šš ÚˋšÝŠŠ Ú´šŠÀ Ú˜ŠŠ¿š õ¡ŠÈ´š§šÊšÊšÀš¡š¯(G6P)š¥ŠÀ Š°ÚšÚ´ŠÊ.
Š´Š¯ÝšÏ ÚÊŠšš
Š´Š¯ÝšÏÚÊŠšš (ÒÓ§Ò°ˆKinase)Š Š´Š¯ÝšÏš š¡š¯ÚÚŠ Ú´šš ÚŠšÇŠÊ. ATP š õ¯ŠÏ šš¿š š¡š¯õ¡¯ŠË¥ Š¥šÇ ŠÇšÇ Š´Š¯ÝšÏš ŠÑš¯ˋšÚ´ŠÊ. š¡ŠÑ šõñ¿š š¡Ú˜ ŠÇŠÀ š Š˜ÚŠ Š¯ šÊšÚ šÙÚ š ÚŠÊ.
ã Siebold, C; Arnold, I; Garcia-Alles, LF; Baumann, U; Erni, B (2003Š
11š 28š¥). “Crystal structure of the Citrobacter freundii dihydroxyacetone kinase reveals an eight-stranded alpha-helical barrel ATP-binding domain.”. The Journal of biological chemistry (ššÇ) 278 (48): 48236ã44. PMID 12966101 . ã Krebs, EG (1983Š
7š 5š¥). “Historical perspectives on protein phosphorylation and a classification system for protein kinases.”. Philosophical transactions of the Royal Society of London. Series B, Biological sciences (ššÇ) 302 (1108): 3ã11. PMID 6137005 . ã Linn, TC (1969Š
9š). “Alpha-keto acid dehydrogenase complexes. XI. Comparative studies of regulatory properties of the pyruvate dehydrogenase complexes from kidney, heart, and liver mitochondria.”. Proc Natl Acad Sci U S A (ššÇ) 64 (1): 227ã234. PMID 6137005 . ã Langan, TA (1969Š
12š). “Phosphorylation of liver histone following the administration of glucagon and insulin.”. Proc Natl Acad Sci U S A (ššÇ) 64 (4): 1276ã83. PMID 4318347 . ã Scheeff, Eric D.; Bourne, Philip E. (2005). “Structural Evolution of the Protein KinaseãLike Superfamily”. PLoS Computational Biology (ššÇ) 1 (5): e49. doi :10.1371/journal.pcbi.0010049 . ã õ¯ Š Manning, G; Whyte, DB; Martinez, R; Hunter, T; Sudarsanam, S (2002Š
12š 6š¥). “The protein kinase complement of the human genome.”. Science (New York, N.Y.) (ššÇ) 298 (5600): 1912ã34. PMID 12471243 . ã õ¯ Š Krebs, EG; Tan, ST; Carrow, DJ; Watts, MK (1985Š
10š). “The phosphorylation of proteins: a major mechanism for biological regulation. Fourteenth Sir Frederick Gowland Hopkins memorial lecture.”. Biochemical Society transactions (ššÇ) 13 (5): 813ã20. PMID 2998902 . ã Samarasinghe, Buddhini. “Hallmarks of Cancer 1:Self-Sufficiency in Growth Signals” . 《Scientific American》. ã Lahiry, Piya; Torkamani, Ali; Schork, Nicholas J.; Hegele, Robert A. (2010Š
1š). “Kinase mutations in human disease: interpreting genotypeãphenotype relationships”. Nature Reviews Genetics (ššÇ) 11 (1): 60ã74. doi :10.1038/nrg2707 . ã Harper, J. W.; Adams, P. D. (2001Š
8š). “Cyclin-Dependent Kinases”. Chemical Reviews (ššÇ) 101 (8): 2511ã2526. doi :10.1021/cr0001030 . ã Karp, Gerald (2010). Cell and molecular biology : concepts and experiments ISBN 9780470483374 ã Lim, S.; Kaldis, P. (2013Š
7š 16š¥). “Cdks, cyclins and CKIs: roles beyond cell cycle regulation”. Development (ššÇ) 140 (15): 3079ã3093. doi :10.1242/dev.091744 . ã õ¯ Š Canavese, Miriam; Santo, Loredana; Raje, Noopur (2012Š
5š 1š¥). “Cyclin dependent kinases in cancer: Potential for therapeutic intervention”. Cancer Biology & Therapy (ššÇ) 13 (7): 451ã457. doi :10.4161/cbt.19589 . ã õ¯ Š Garrington, TP; Johnson, GL (1999Š
4š). “Organization and regulation of mitogen-activated protein kinase signaling pathways.”. Current opinion in cell biology (ššÇ) 11 (2): 211ã8. PMID 10209154 . ã Sun, Yue; Thapa, Narendra; Hedman, Andrew C.; Anderson, Richard A. (2013Š
6š). “Phosphatidylinositol 4,5-bisphosphate: Targeted production and signaling”. BioEssays (ššÇ) 35 (6): 513ã522. doi :10.1002/bies.201200171 . ã Heath, CM; š¡. (2003). “Lipid Kinases Play Crucial and Multiple Roles in Membrane Trafficking and Signalling” (PDF) . Histology and Histopathology (ššÇ) 18 : 989ã998. ã Cantley, Lewis C (2012). “PI 3-kinase and disease”. BMC Proceedings (ššÇ) 6 (Suppl 3): O2. doi :10.1186/1753-6561-6-S3-O2 . ã õ¯ Š ŠÊ Miller, S.; Tavshanjian, B.; Oleksy, A.; Perisic, O.; Houseman, B. T.; Shokat, K. M.; Williams, R. L. (2010Š
3š 25š¥). “Shaping Development of Autophagy Inhibitors with the Structure of the Lipid Kinase Vps34”. Science (ššÇ) 327 (5973): 1638ã1642. doi :10.1126/science.1184429 . ã Neubauer, Heidi A.; Pitson, Stuart M. (2013Š
11š). “Roles, regulation and inhibitors of sphingosine kinase 2”. 《FEBS Journal》 280 (21): 5317ã5336. doi :10.1111/febs.12314 . ã Holzer, H; Duntze, W (1971). “Metabolic regulation by chemical modification of enzymes.”. Annual review of biochemistry (ššÇ) 40 : 345ã74. PMID 4399446 . ã “Nonspherocytic hemolytic anemia due to hexokinase deficiency” . 2015Š
9š 5š¥š šŠ°¡ Š˜¡š šš Š°ÇšÀÇŠ Š˜¡š. 2014Š
6š 22š¥š Úš¡Ú´ . ã “Phosphofructokinase Deficiency Glycogen Storage Disease” . 2015Š
4š 19š¥š šŠ°¡ Š˜¡š šš Š°ÇšÀÇŠ Š˜¡š. 2014Š
6š 22š¥š Úš¡Ú´ . ã Bauer, S; Kemter, K; Bacher, A; Huber, R; Fischer, M; Steinbacher, S (2003Š
3š 7š¥). “Crystal structure of Schizosaccharomyces pombe riboflavin kinase reveals a novel ATP and riboflavin-binding fold.”. Journal of molecular biology (ššÇ) 326 (5): 1463ã73. PMID 12595258 . ã Pratt, Donald Voet, Judith G. Voet, Charlotte W. (2008). Fundamentals of biochemistry : life at the molecular level (ššÇ) 3Ú. Hoboken, NJ: Wiley. ISBN 9780470129302 ã õ¯ Š Karthikeyan, S; Zhou, Q; Osterman, AL; Zhang, H (2003Š
11š 4š¥). “Ligand binding-induced conformational changes in riboflavin kinase: structural basis for the ordered mechanism.”. Biochemistry (ššÇ) 42 (43): 12532ã8. PMID 14580199 . ã Galluccio, M; Brizio, C; Torchetti, EM; Ferranti, P; Gianazza, E; Indiveri, C; Barile, M (2007Š
3š). “Over-expression in Escherichia coli, purification and characterization of isoform 2 of human FAD synthetase.”. Protein expression and purification (ššÇ) 52 (1): 175ã81. PMID 17049878 . ã Zou, YX; Zhang, XH; Su, FY; Liu, X (2012Š
10š). “Importance of riboflavin kinase in the pathogenesis of stroke.”. CNS neuroscience & therapeutics (ššÇ) 18 (10): 834ã40. PMID 22925047 . ã Brijlal, Sangeetha; Lakshmi, A. V; Bamji, Mahtab S.; Suresh, P. (2007Š
3š 9š¥). “Flavin metabolism during respiratory infection in mice”. British Journal of Nutrition (ššÇ) 76 (03): 453. doi :10.1079/BJN19960050 . ã Aufderklamm, S; TodenhûÑfer, T; Gakis, G; Kruck, S; Hennenlotter, J; Stenzl, A; Schwentner, C (2012Š
3š). “Thymidine kinase and cancer monitoring.”. Cancer letters (ššÇ) 316 (1): 6ã10. PMID 22068047 . ã Topolcan, Ondrej; Holubec, Lubos (2008Š
2š). “The role of thymidine kinase in cancer diseases”. Expert Opinion on Medical Diagnostics (ššÇ) 2 (2): 129ã141. doi :10.1517/17530059.2.2.129 . ã Gotz, A.; Isohanni, P.; Pihko, H.; Paetau, A.; Herva, R.; Saarenpaa-Heikkila, O.; Valanne, L.; Marjavaara, S.; Suomalainen, A. (2008Š
6š 21š¥). “Thymidine kinase 2 defects can cause multi-tissue mtDNA depletion syndrome”. Brain (ššÇ) 131 (11): 2841ã2850. doi :10.1093/brain/awn236 .