Masrasector ist eine Gattung aus der Ordnung der Hyaenodonta, ausgestorbenen fleischfressenden Säugetieren, die möglicherweise den Raubtieren nahe stehen. Sie kam im Oberen Eozän und im Unteren Oligozän vor 38 bis 30 Millionen Jahren im heutigen Afrika und auf der Arabischen Halbinsel vor. Das Fundmaterial setzt sich hauptsächlich aus Schädel- und Gebissresten zusammen, darüber hinaus liegen auch einzelne Elemente des Bewegungsapparates vor. Als bedeutendster Fundpunkt erwies sich die Fossillagerstätte des Fayyum in Ägypten. Anhand des bekannten Fossilmaterials kann auf kleine Tiere geschlossen werden. Möglicherweise ernährten sie sich wiederum von kleineren Säugetieren, es wurde aber auch eine Spezialisierung auf Weichtieren in Betracht gezogen. Die wissenschaftliche Einführung der Gattung datiert in das Jahr 1974, gegenwärtig sind mehrere Arten belegt. Anfangs war die systematische Stellung von Masrasector aufgrund einiger besonderer Zahnmerkmale umstritten.
Masrasector war ein eher kleiner Vertreter der Hyaenodonta. Kleine Angehörige der Gattung wiesen ein rekonstruiertes Gewicht von durchschnittlich 1,05 bis 1,16 kg auf. Größere Exemplare erreichten nicht die Ausmaße des nahe verwandten Brychotherium. In der Größe entsprachen die Tiere somit der heutigen Kleinfleck-Ginsterkatze oder dem Streifenskunk. Von der Gattung liegen der Schädel und einzelne Teile des Bewegungsapparates vor. Die bekannten Schädel sind vollständig bei einer Länge von 8,1 cm, sie zeigen aber markante lagerungsbedingte Deformationen. Das Rostrum war schmal und teils röhrenförmig. Der Mittelkieferknochen bestand nur als dünner Knochen, der den seitlichen Rand des Naseninnenraumes bildete. Das Nasenbein zog über der Nasenöffnung seitlich etwas ein. Die Sutur zwischen dem Nasen- und dem Stirnbein bildete ein V mit der Spitze nach hinten weisend. Am Oberkiefer lag oberhalb des dritten Prämolaren ein großes Foramen infraorbitale, das einen Durchmesser von 3,2 mm besaß. Der Oberkiefer selbst war nicht am Rand der Orbita beteiligt. Dieser wurde oben vom Stirnbein, vorn vom Tränenbein und unten vom Schläfenbein gebildet, der vordere Rand saß oberhalb des ersten Molaren. Das Stirnbein zeigte am oberen hinteren Rand der Orbita eine markante Aufwölbung, ein Processus postorbitalis war aber nicht ausgebildet. Das Tränenbein ragte von der Orbita ins Gesicht und besaß einen weiten Kontaktbereich mit dem Oberkiefer. Der Jochbogen wies über seine gesamte Länge einen recht massiven Bau auf. Zwei markante Temporallinien verliefen über das Stirnbein und dienten als Ansatz des Musculus temporalis. Auf dem Scheitelbein war zusätzlich ein kräftiger Scheitelkamm ausgebildet, der bis zum Wulst des Hinterhauptsbeins reichte. Dieser war fächerartig breit und ausgedehnt. Die Gelenkflächen des Hinterhauptsbeins zum Anschluss an die Halswirbelsäule hatten eine ovale Form und waren so höher als breit. Die Schädelbasis ist weitgehend beschädigt. Die Glenoidgrube zur Gelenkung des Unterkiefers mit dem Schädel war ebenfalls oval mit der größten Breite quer zur Schädelachse.[1]
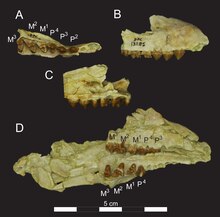
Verschiedene Unterkiefer von Masrasector in jeweils drei Ansichten
Der Unterkiefer besaß einen massiven Bau. Die Symphyse reichte bis zum dritten Prämolaren und war nicht geschlossen. Das vordere Foramen mentale befand sich unter dem ersten Prämolaren, zwei weitere folgten jeweils unter dem dritten und vierten Vormahlzahn. Der untere Rand des Unterkiefers verlief konvex bis etwa zur Mitte unter dem Kronenfortsatz. Hier war eine kleine konkave Einbuchtung ausgebildet, die dann zum gerundeten Winkelfortsatz auslief. Unterhalb der Spitze des Kronenfortsatzes öffnete sich das Foramen mandibulae. Es war horizontal orientiert, der dort ansetzende Mandibularkanal maß 2 mm im Durchmesser. Der aufsteigende Ast des Unterkiefers setzte in einem Winkel von 106° zur Alveolarebene an. Er war breit gestaltet und ragte hoch auf, etwa um das 1,25fache der Höhe des horizontalen Knochenkörpers.[1]
Verschiedene Oberarmfragmente von Masrasector in jeweils drei Ansichten
Weder in der oberen noch in der unteren Gebisshälfte sind die Schneidezähne erhalten. Der Eckzahn repräsentierte den größten Zahn, im Unterkiefer erreichte er die Hälfte der Höhe des Kronenfortsatzes. Er hatte eine seitlich gepresste Gestalt und war leicht nach hinten gekrümmt. Es waren keine scharfen Schneidkanten ausgebildet, so dass der Querschnitt eher konisch anstatt klingenartig wirkte. Alle hinteren Zähne standen in einer geschlossenen Reihe. Die Prämolaren wiesen in der Seitenansicht eine massiv dreieckige Gestalt auf, was vor allem durch den großen Paraconus (Oberkiefer) beziehungsweise das Protoconid (Unterkiefer) hervorgerufen wurde, den jeweiligen Haupthöcker der Vormahlzähne. Bei Masrasector war der Höcker aber eher buckelig geformt und nicht so spitz wie beim verwandten Brychotherium. Auch fehlten den hintersten Prämolaren weitgehend die scharfen Schneidkanten, die etwa bei Brychotherium bestanden. Die Molaren besaßen einen typisch sectorialen Bau mit drei Haupthöckern (Metaconus, Paraconus und Protoconus bei den oberen sowie Metaconid, Paraconid und Protoconid bei den unteren Mahlzähnen). Der Meta- und der Paraconus der ersten beiden Oberkiefermolaren waren seitlich etwas gepresst, jedoch nicht so stark wie bei Brychotherium. Die Höcker bildeten an der Basis eine Einheit, die Spitzen standen deutlich auseinander. Auf dem ersten Molar ragten beide Höcker etwa gleich hoch, auf dem zweiten war der Metaconus leicht höher als der Paraconus. Die an der Basis fusionierten Höcker und der gegenüber dem Paraconus höhere Metaconus sind charakteristische Kennzeichen der Teratodontinae, bei stammesgeschichtlich entwickelten Formen wie Dissopsalis und Anasinopa war der Paraconus zudem deutlich in der Größe reduziert. An den seitlichen Kanten bestanden scharfe Scherleisten, wobei die hintere (Postmetacrista) über eine flache Bucht mit dem Metastyl in Verbindung stand, die Bucht war auf dem zweiten Molaren markanter als auf dem ersten. Der Protoconus ragte etwa halb so hoch wie Meta- und Paraconus. Der dritte Oberkiefermolar war deutlich kleiner als die beiden vorherigen und zeichnete sich durch das Fehlen des Metaconus aus. Im Unterkiefer wiesen die Molaren ein charakteristisches Talonid auf (einen niedriger liegenden Teil der Kauoberfläche), das knapp die Hälfte der Zahnlänge beanspruchte. Dadurch wich Masrasector von Brychotherium mit seinen kürzeren Talonids ab und entsprach eher Glibzegdouia. Den größten Höcker bildete das Protoconid, den kleinsten das Metaconid, während das Paraconid zwischen beiden vermittelte. Vom vordersten zum hintersten Unterkiefermolar wuchs das Paraconid verhältnismäßig an Größe, das Metaconid wurde dagegen kleiner. In diesem Merkmal stimmt Masrasector mit Brychotherium überein. Der größte Zahn im Unterkiefer war der hinterste Molar mit 6,2 mm Länge. Teilweise übertrafen die Prämolaren aber die vorderen Molaren an Länge. So war der zweite Prämolar 5,4 mm lang, der erste Molar aber nur 4,5 und der zweite 5,3 mm. Ähnlich verhielt es sich im Oberkiefer, hier betrugen die entsprechenden Werte für den zweiten Prämolaren 5,2 mm (der erste ist unbekannt), für den ersten Molaren 4,7 und für den zweiten Molaren 5,9 mm. Der wesentlich kleinere dritte Molar erreichte nur 2,7 mm Länge.[1]
Es sind lediglich drei Fragmente des unteren Oberarmknochens bekannt. Das längste Stück ist 63 mm lang und besitzt noch substantielle Teile der Diaphyse. Der Schaft war rundlich im Querschnitt und wies eine markante Knochenleiste als Ansatzpunkt des Musculus supinator auf. Die Leiste verschmälerte sich zum unteren Gelenkende (Ellenbogengelenk) hin und lief dann auf der Außenseite auf dem äußeren Vorsprung (Epicondyle) der Gelenkrolle aus. Der innere Vorsprung der Gelenkrolle war vergrößert. Die Fossa olecrani bildete oberhalb der Gelenkrolle eine massive Öffnung im Knochen.[1]
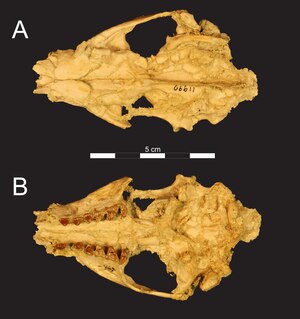
Fossilfunde
Oberkieferfragment (A–C) und Schnauzenpartie (D) von Masrasector
Masrasector ist von mehreren Fundstellen in Nordafrika und der Arabischen Halbinsel bekannt. Zu den umfangreichsten Fossilresten gehören die der Fayyum-Region im nördlichen Ägypten, eine der bedeutendsten Fossillagerstätten aus dem Paläogen. Das bisher bekannte Material streut stratigraphisch relativ weit. Die möglicherweise ältesten Funde stammen hier noch aus der Fundlokalität BQ-2 im unteren Abschnitt der Qasr-el-Sagha-Formation und gehören damit in das Obere Eozän vor etwa 37 Millionen Jahren. Es handelt sich um einzelne isolierte Zähne und einen Unterkieferast.[2] Sehr umfangreich sind die Fossilfunde aus der Lokalität L-41, die sich im untersten Abschnitt der Gebel-Qatrani-Formation befindet. Das Alter beträgt etwa 34 Millionen Jahre. Die hier sehr feinkörnigen Ablagerungen unterscheiden sich von den restlichen gröberklastischen Sedimenten der Gesteinseinheit und gehen wohl auf einen ehemaligen See zurück. Bisher entdeckt wurden hier mehrere vollständige Schädel und zahlreiche Unterkiefer sowie einzelne Langknochen. Damit gehört Masrasector zu den kleinsten Hyaenodonten des afroarabischen Raumes, von denen Teile des Bewegungsapparates überliefert sind.[1] Aus jüngeren Schichten der Gebel-Qatrani-Formation, die weitgehend in das Untere Oligozän datieren, sind einige Einzelfunde belegt. Hervorzuheben sind ein Unterkieferrest, ein Oberkieferfragment und isolierte Zähne aus der Fundstelle G. Sie stellen das forschungsgeschichtlich zuerst entdeckte Fundmaterial dar und dienten zur Aufstellung der Gattung.[3] Weitere dokumentierte Funde von Masrasector erhielten sich an den Fundstellen A, E, I und M. Jene der beiden letztgenannten stellen die jüngsten Funde der Gattung im Fayyum dar.[4]
Abseits der Fayyum-Region wurde Masrasector in Nordafrika in Bir el-Ater (Nementcha) im nordöstlichen Algerien aufgefunden. Die Fundstelle mit ihren reichhaltigen Fossilresten gehört in das Obere Eozän, genauer in den Übergang von Bartonium zu Priabonium. Dadurch bilden die Funde von Masrasector hier wohl die ältesten der Gattung bisher. Sie beschränken sich aber lediglich auf zwei Ober- und einen Unterkieferbackenzahn.[4] Zwei weitere Zähne kamen aus der Ashawq-Formation bei Taqah im südlichen Oman zum Vorschein. Sie haben ein unteroligozänes Alter und dürften daher zeitgleich zu den Resten aus dem oberen Bereich der Gebel-Qatrani-Formation im Fayyum sein.[5]
Paläobiologie
Anhand des unteren Gelenkendes des Oberarmknochens lässt sich ein stabiles Ellenbogengelenk annehmen. Es kann daraus geschlussfolgert werden, dass Masrasector sich wohl terrestrisch fortbewegte, möglicherweise war er ein schneller Läufer. Generell haben Hyaenodonten aber einen massiveren Bewegungsapparat als heutige Raubtiere. Ausgehend von dem geringen Körpergewicht der Tiere lässt sich ihr Beutespektrum im Fayyum auf die kleineren stachelschweinartigen Nagetiere und die kleinen Vertreter der Schliefer einengen. Die eher baumbewohnenden Primaten sind wahrscheinlich als potentielle Nahrungsressource auszuschließen.[1] Andererseits könnten die eher stumpfen Höcker der Prämolaren auch auf eine Bevorzugung von hartschaligen Mollusken hindeuten (durophag).[6]
Systematik
Innere Systematik der Teratodontinae nach Solé & Mennecart 2019[7]
Masrasector ist eine Gattung aus der ausgestorbenen Unterfamilie der Teratodontinae innerhalb der ebenfalls erloschenen Ordnung der Hyaenodonta. Die Hyaenodonta galten ursprünglich als Teil der Creodonta, die teilweise etwas irreführend auch als „Urraubtiere“ bezeichnet werden. Es wurde angenommen, dass die Creodonta die Schwestergruppe der heutigen Raubtiere (Carnivora) innerhalb der übergeordneten Gruppe der Ferae darstellen.[8] Da sich die Creodonta in der Folgezeit als in sich nicht geschlossene Gruppe erwiesen, wurden sie in die Hyaenodonta und die Oxyaenodonta aufgespalten.[9][10] Beide Gruppen zeichnen sich gegenüber den Raubtieren durch eine weiter nach hinten im Gebiss verlagerte Brechschere aus. In der Regel sind bei den Hyaenodonten der zweite Oberkiefer- und der dritte Unterkiefermolar einbezogen. Die Hyaenodonten lassen sich erstmals im Mittleren Paläozän vor rund 60 Millionen Jahren nachweisen, ihr letztes Auftreten haben sie im Mittleren Miozän vor etwa 9 bis 10 Millionen Jahren. Die Teratodontinae nehmen innerhalb der Hyaenodonta die Position der Schwestergruppe der Familie der Hyainailouridae ein, beide zusammen formen die übergeordnete Gruppe der Hyainailouroidea. Ein charakteristisches Merkmal der Teratodontinae findet sich im Bau der Oberkiefermolaren, bei denen der Para- und der Metaconus nur an der Basis miteinander fusioniert sind und letzterer ersteren überragt. Im Unterschied dazu verschmelzen bei den Hyainailouridae der Para- und der Metaconus zum Amphiconus. Außerdem ist der Paraconus höher als der Metaconus. In Bezug auf die Höhenausprägung der beiden Höcker stimmen die Teratodontinae mit den Hyaenodontidae überein, bei diesen sind der Para- und der Metaconus aber ebenfalls miteinander verwachsen. Masrasector bildet eine nähere Verwandtschaftsgruppe mit einigen stammesgeschichtlich jüngeren Vertretern der Teratodontinae, etwa Dissopsalis, Teratodon und Anasinopa. Als Schwestergattung der Gruppe kann Brychotherium aufgefasst werden, das ebenfalls im Fayyum nachgewiesen ist.[11][12]
Folgende Arten von Masrasector sind anerkannt:[3][5][1]
Von diesen drei Arten ist M. nananubis der stammesgeschichtlich älteste und kleinste Vertreter, er trat bereits im Oberen Eozän an der Fundstelle L-41 in Fayyum auf.[1] Die beiden anderen Arten datieren weitgehend in das Untere Oligozän. M. aegypticum stammt aus der Fundstelle G des Fayyum.[3]M. ligabuei wurde zuerst in Taqah in Oman entdeckt,[5] die Funde aus Bir el-Ater könnten aber eventuell ebenfalls diese Art repräsentieren, womit der Vertreter ebenfalls bereits im Oberen Eozän in Erscheinung trat.[4] Eine potentielle weitere Art, „M. pithecodacos“, war bereits 1994 in einer unveröffentlichten Arbeit von Patricia A. Holroyd für die Fundstellen I und M des Fayyum-Gebietes aufgestellt worden, eine formal gültige Beschreibung liegt aber bisher nicht vor.[4] Ein kleinerer Unterkiefer aus der Fundstelle G im Fayyum wurde zunächst als mögliche neue Art gedeutet,[3] später aber als Jungtier von M. aegypticum interpretiert; anderen Meinungen zufolge repräsentiert er eine bisher nicht beschriebene Gattung.[6]
Masrasector wurde im Jahr 1974 von Elwyn L. Simons und Philip D. Gingerich wissenschaftlich erstbeschrieben. Das Typusmaterial stammt aus der Fundstelle G der Fayyum-Region in Ägypten. Der Holotyp (Exemplarnummer CGM 30978) besteht aus einem linken Unterkiefer mit dem letzten Prämolaren und den drei Molaren. Der Gattungsname Masrasector setzt sich aus dem arabischen Wort Masr für „Ägypten“ und dem lateinischen Wort sector für „Zerschneider“ (von secare „zerschneiden“) zusammen. In ihrer Erstbeschreibung sahen die Autoren Masrasector noch in der Nähe von ursprünglichen Hyaenodonten wie Proviverra, Sinopa und Metasinopa mit einer intermediären Stellung zwischen den beiden letztgenannten. Sie begründeten dies mit der ihrer Meinung nach progressiven Vergrößerung der drei Haupthöcker der Unterkiefermolaren und der zunehmenden Bedeutung der Schneidkanten.[3] In der nachfolgenden Zeit wurde Masrasector gemeinsam mit anderen Formen wie Anasinopa und Dissopsalis daher immer wieder als „afrikanische Proviverrinen“ angesehen, die genauen verwandtschaftlichen Beziehungen blieben weitgehend unklar. Je nach Wichtung der Zahnmerkmale vermuteten Wissenschaftler einerseits eine engere Bindung an ähnliche Formen aus Südostasien (Kyawdawia und Paratritemnodon),[14][15] andererseits auch eine Nahverwandtschaft mit einigen amerikanischen Formen (Sinopa und Prototomus).[16] Die eher stumpfe und große Ausprägung der Prämolaren ließ wiederum einige Forscher über eine nähere Verwandtschaft mit dem aus dem Miozän Afrikas bekannten Teratodon spekulieren, das bisher nur spärlich belegt ist.[6]Phylogenetische Untersuchungen bestätigten diese Ansicht später und führten somit zu einem Verweis zu den Teratodontinae.[17][18][12][1]
Literatur
Matthew R. Borths und Erik R. Seiffert: Craniodental and humeral morphology of a new species of Masrasector (Teratodontinae, Hyaenodonta, Placentalia) from the late Eocene of Egypt and locomotor diversity in hyaenodonts. PLoS ONE 12 (4), 2017, S. e0173527, doi:10.1371/journal.pone.0173527
Elwyn L. Simons und Philip D. Gingerich: New carnivorous mammals from the Oligocene of Egypt. Annals of the Geological Survey of Egypt 4, 1974, S. 157–166
Einzelnachweise
↑ abcdefghiMatthew R. Borths und Erik R. Seiffert: Craniodental and humeral morphology of a new species of Masrasector (Teratodontinae, Hyaenodonta, Placentalia) from the late Eocene of Egypt and locomotor diversity in hyaenodonts. PLoS ONE 12 (4), 2017, S. e0173527, doi:10.1371/journal.pone.0173527
↑Matthew R. Borths, Erik R. Seiffert, Katherine Goodenberger und Elwyn L. Simons: The oldest Fayum Creodont: Dental and Humeral morphology of a new proviverrine hyaenodontid from the earliest Late Eocene of Egypt. Journal of Vertebrate Paleontology 30 (suppl), 2010, S. 63A
↑ abcdeElwyn L. Simons und Philip D. Gingerich: New carnivorous mammals from the Oligocene of Egypt. Annals of the Geological Survey of Egypt 4, 1974, S. 157–166
↑ abcdFloréal Solé, El Mabrouk Essid, Wissem Marzougui, Rim Temani, Hayet Khayati Ammar, Mhammed Mahboubi, Laurent Marivaux, Monique Vianey-Liaud und Rodolphe Tabuce: New fossils of Hyaenodonta (Mammalia) from the Eocene localities of Chambi (Tunisia) and Bir el Ater (Algeria), and the evolution of the earliest African hyaenodonts. Palaeontologia Electronica 19 (3), 2016, S. 41A ([1])
↑ abcJean-Yves Crochet, Herbert Thomas Jack Roger, Sevket Sen und Zaher Al-Sulaimani: Première découverte d'unc créodonte dans la péninsule Arabique: Masrasector ligabuei nov. sp. (Oligocène inférieur deTaqah, Formation d'Ashawq, Sultanat d'Oman). Comptes rendus de l'Académie des sciences. Série 2 311, 1990, S. 1455–1460 ([2])
↑ abcMargaret E. Lewis und Michael Morlo: Creodonta. In: Lars Werdelin und William Joseph Sanders (Hrsg.): Cenozoic Mammals of Africa. University of California Press, Berkeley, Los Angeles, London, 2010, S. 543–560
↑Floréal Solé und Bastien Mennecart: A large hyaenodont from the Lutetian of Switzerland expands the body mass range of the European mammalian predators during the Eocene. Acta Palaeontologica Polonica 64, 2019, doi:10.4202/app.00581.2018
↑Kenneth D. Rose: The beginning of the age of mammals. Johns Hopkins University Press, Baltimore, 2006, S. 1–431 (S. 122–126)
↑Michael Morlo, Gregg Gunnell und P. David Polly: What, if not nothing, is a creodont? Phylogeny and classification of Hyaenodontida and other former creodonts. Journal of Vertebrate Paleontology 29 (3 suppl), 2009, S. 152A
↑Floréal Solé: New proviverrine genus from the Early Eocene of Europe and the first phylogeny of Late Paleocene-Middle Eocene hyaenodontidans (Mammalia). Journal of Systematic Paleontology 11, 2013, S. 375–398
↑Floréal Solé, Eli Amson, Matthew Borths, Dominique Vidalenc, Michael Morlo und Katharina Bastl: A New Large Hyainailourine from the Bartonian of Europe and Its Bearings on the Evolution and Ecology of Massive Hyaenodonts (Mammalia). PLoS ONE 10 (9), 2015, S. e0135698, doi:10.1371/journal.pone.0135698
↑ abMatthew R. Borths, Patricia A. Holroyd und Erik R. Seiffert: Hyainailourine and teratodontine cranial material from the late Eocene of Egypt and the application of parsimony and Bayesian methods to the phylogeny and biogeography of Hyaenodonta (Placentalia, Mammalia). PeerJ 4, 2016, S. e2639 doi:10.7717/peerj.2639
↑Matthew R. Borths und Nancy J. Stevens: The first hyaenodont from the late Oligocene Nsungwe Formation of Tanzania: Paleoecological insights into the Paleogene-Neogene carnivore transition. PLoS ONE 12 (10), 2017, S. e0185301, doi:10.1371/journal.pone.0185301
↑Naoko Egi, Patricia A. Holroyd, Takeshi Tsubamoto, Nobuo Shigehara, Masanaru Takai, Soe Thura Tun, Aye Ko Aung und Aung Naing Soe: A new genus and species of hyaenodontid creodont from the Pondaung Formation (Eocene, Myanmar). Journal of Vertebrate Paleontology 24 (2), 2004, S. 502–506
↑Naoko Egi, Patricia A. Holroyd, Takehisa Tsubamoto, Aung Naing Soe, Masanaru Takai und Russell L. Ciochon: Proviverrine hyaenodontids (Creodonta: Mammalia) from the Eocene of Myanmar and a phylogenetic analysis of the proviverrines from the para-Tethys area. Journal of Systematic Palaeontology 3 (4), 2005, S. 337–358
↑Stéphane Peigné, Michael Morlo, Yaowalak Chaimanee, Stéphane Durocq, Soe Thura Tun und Jean-Jacques Jaeger: New discoveries of hyaenodontids (Creodonta, Mammalia) from the Pondaung Formation, middle Eocene, Myanmar – paleobiogeographic implications. Geodiversitas 29 (3), 2004, S. 441–458
↑Floréal Solé, Julie Lhuillier, Mohammed Adaci, Mustapha Bensalah, Mohammed Mahboubi und Rodolphe Tabuce: The hyaenodontidans from the Gour Lazib area (?Early Eocene, Algeria): implications concerning the systematics and the origin of the Hyainailourinae and Teratodontinae. Journal of Vertebrate Paleontology 12 (3), 2014, S. 303–322
↑Rajendra S. Rana, Kishor Kumar, Shawn P. Zack, Floréal Solé, Kenneth D. Rose, Pieter Missiaen, Lachham Singh, Ashok Sahni und Thierry Smith: Craniodental and Postcranial Morphology of Indohyaenodon raoifrom the Early Eocene of India, and Its Implications for Ecology, Phylogeny, and Biogeography of Hyaenodontid Mammals. Journal of Vertebrate Paleontology 35 (5), 2015, S. e965308, doi:10.1080/02724634.2015.965308
Neighborhood of Baltimore in Maryland, United StatesMidtown-EdmondsonNeighborhood of BaltimoreMosher St. Swell-Front RowhousesMidtown-EdmondsonLocation in Baltimore, MDCoordinates: 39°17′46″N 76°39′01″W / 39.29611°N 76.65028°W / 39.29611; -76.65028CountryUnited StatesStateMarylandCityBaltimoreTime zoneUTC-5 (Eastern) • Summer (DST)UTC-4 (EDT)ZIP code21217ZIP code21223 Midtown-Edmondson is a mixed-use neighborhood in western Baltimore City develo...
Osservatorio meteorologico di Vienna Hohe WarteStato Austria LandVienna ComuneVienna Altitudine202 m s.l.m. Codice WMO11035 GestoreZAMG T. media gennaio0,1 °C T. media luglio20,2 °C T. media annua10,2 °C T. max. assoluta38,5 °C T. min. assoluta-26,3 °C Prec. medie annue620,3 mm Coordinate48°14′54.55″N 16°21′22.47″E / 48.248486°N 16.356242°E48.248486; 16.356242Coordinate: 48°14′54.55″N 16°21′22.47″E...
A Federação Russa e suas Regiões Econômicas 1. Região Econômica do Centro 2. Região Econômica Central da Terra Negra 3. Região Econômica do Leste Siberiano 4. Região Econômica do Extremo Oriente 5. Região Econômica do Norte 6. Região Econômica do Norte do Cáucaso 7. Região Econômica do Noroeste 8. Região Econômica do Volga 9. Região Econômica dos Urais 10. Região Econômica Volgo-Viatski 11. Região Econômica do Oeste Siberiano 12. Região Econômica de Kaliningrado A...
Pulau Ubi Besar Pulau RotterdamPulau Pulau Ubi Besar dahulu merupakan pulau yang berada pada gugusan Kepulauan Seribu yang secara administratif termasuk dalam wilayah Kabupaten Administratif Kepulauan Seribu provinsi DKI Jakarta saat sekarang pulau ini telah tenggelam akibat eksploitasi pasir laut.[1] Lihat pula Kabupaten Administratif Kepulauan Seribu Kepulauan Seribu Referensi ^ News: Dua Pulau di Kepulauan Seribu Karam. Diarsipkan dari versi asli tanggal 2016-03-05. Diakses tanggal...
Este artículo o sección tiene referencias, pero necesita más para complementar su verificabilidad.Este aviso fue puesto el 24 de junio de 2022. Kōsuke Kimura Kimura en 2011Datos personalesNombre de nacimiento 木村光佑Nacimiento Kōbe, Japón14 de mayo de 1984 (39 años)Nacionalidad(es) JaponesaAltura 1,75 metrosCarrera deportivaDeporte FútbolClub profesionalDebut deportivo 2007(Colorado Rapids)Club Nashville SCLiga USL ChampionshipPosición CentrocampistaDorsal(es) 27[edita...
Coloring BookAlbum mini karya Oh My GirlDirilis03 April 2017 (2017-04-03)Direkamdoobdoob StudioVibe StudioW SoundKoko Sound StudioBono StudioGenreK-popDurasi15:48BahasaKoreaLabelWM EntertainmentProduserLee Won-min72Moon Jeong-gyuKronologi Oh My Girl Listen to My Word(2016)Listen to My Word2016 Coloring Book(2017) Secret Garden(2018)Secret Garden2018 Singel dalam album Coloring Book Coloring BookDirilis: 3 April 2017 Coloring Book (Hangul: 컬러링북) adalah album mini keli...
يفتقر محتوى هذه المقالة إلى الاستشهاد بمصادر. فضلاً، ساهم في تطوير هذه المقالة من خلال إضافة مصادر موثوق بها. أي معلومات غير موثقة يمكن التشكيك بها وإزالتها. (يونيو_2011) ترتيب المدارك وتقريب المسالك لمعرفة أعلام مذهب مالكمعلومات عامةالمؤلف القاضي عياض اللغة العربية الموضوع...
Lage der Gemarkung Gompitz in Dresden Die Liste der Straßen und Plätze in Gompitz beschreibt das Straßensystem im Dresdner Ortsteil Gompitz mit den entsprechenden historischen Bezügen. Aufgeführt sind Straßen und Plätze, die im Gebiet der Gemarkung Gompitz liegen. Kulturdenkmale in der Gemarkung Gompitz sind in der Liste der Kulturdenkmale in Gompitz aufgeführt. Das Gebiet von Gompitz ist seit 1999 der gleichnamigen Ortschaft der sächsischen Landeshauptstadt Dresden zugewiesen. Insge...
13th quadrennial U.S. presidential election 1836 United States presidential election ← 1832 November 3 – December 7, 1836 1840 → 294 members of the Electoral College148 electoral votes needed to winTurnout56.5%[1] 0.5 pp Nominee Martin Van Buren William Henry Harrison Hugh L. White Party Democratic Whig Whig Alliance Anti-Masonic Home state New York Ohio Tennessee Running mate Richard M. Johnson Francis Granger John Tyler Electoral vote 17...
This article does not cite any sources. Please help improve this article by adding citations to reliable sources. Unsourced material may be challenged and removed.Find sources: Maveli Stores – news · newspapers · books · scholar · JSTOR (January 2012) (Learn how and when to remove this template message) Logo of Maveli Stores Maveli Stores is a venture of the Kerala State Civil Supplies Corporation (Supplyco). Under the Government programme of market in...
For information on all Boston University sports, see Boston University Terriers. College football program of Boston University Boston University Terriers footballFirst season1884[1][2]Last season1997StadiumNickerson Field(capacity: 10,412)Field surfaceFieldTurfLocationBoston, MassachusettsNCAA divisionDivision I-AAConferenceAtlantic 10 ConferenceAll-time record323–390–34 (.455)Bowl record0–1 (.000)Conference titles5 (1980, 1982, 1983, 1984, 1993)Riv...
Blessing of the Fleet event logo Blessing of the Fleet procession, Port Adelaide, South Australia, 2015 The Blessing of the Fleet is a tradition that began centuries ago in Mediterranean fishing communities. The practice began predominantly Catholic, but is now practiced by all Christians as a blessing from the local priest and pastors that is meant to ensure a safe and bountiful season. In most ports, the event was brought by immigrants who held strongly to their Christian religious beliefs....
Nonfiction book by poet and critic Maggie Nelson The Argonauts AuthorMaggie NelsonCountryUnited StatesGenreAutotheoryPublisherGraywolf PressPublication date2015Media typePrintPages160ISBN1555977073 This article is about the book by Maggie Nelson. For the mythological heroes, see Argonauts. For other uses, see Argonaut (disambiguation). The Argonauts is a book by poet and critic Maggie Nelson, published in 2015. It mixes philosophical theory with memoir.[1] The book discusses her ...
هذه المقالة يتيمة إذ تصل إليها مقالات أخرى قليلة جدًا. فضلًا، ساعد بإضافة وصلة إليها في مقالات متعلقة بها. (يوليو 2023) عمر فاروق عثمان (صحفي) معلومات شخصية مواطنة الصومال الحياة العملية المهنة صحفي اللغات الصومالية، والإنجليزية بوابة إعلام تعديل مصدري - تعدي...
Castle of MytileneΚάστρο της ΜυτιλήνηςLesbos, North Aegean Castle of Mytilene (2010)Castle of MytileneCoordinates39°38′35″N 26°20′04″E / 39.643°N 26.3345°E / 39.643; 26.3345Typehilltop citadelSite informationOwnerGreek Ministry of CultureOpen tothe publicYesConditionruinSite historyBuilt byByzantine EmpireMaterialshewn stone (ashlar) The Castle of Mytilene, also Fortress of Mytilene, is located in Mytilene on the Greek islan...
Species of mammal White-nosed coati at Tikal, Guatemala Conservation status Least Concern (IUCN 3.1)[1] Scientific classification Domain: Eukaryota Kingdom: Animalia Phylum: Chordata Class: Mammalia Order: Carnivora Family: Procyonidae Genus: Nasua Species: N. narica Binomial name Nasua narica(Linnaeus, 1766) Subspecies[2] N. n. narica (Linnaeus, 1766) N. n. molaris Merriam, 1902 N. n. nelsoni Merriam, 1901 N. n. yucatanica J. A. Allen, 1904 The native range of ...
Japanese manga series You and I Are Polar OppositesFirst tankōbon volume cover正反対な君と僕(Seihantai na Kimi to Boku)GenreRomantic comedy[1] MangaWritten byKōcha AgasawaPublished byShueishaEnglish publisherNA: Viz MediaImprintJump Comics+MagazineShōnen Jump+DemographicShōnenOriginal runMay 2, 2022 – presentVolumes5 You and I Are Polar Opposites (Japanese: 正反対な君と僕, Hepburn: Seihantai na Kimi to Boku) is a Japanese manga series written and illustra...










")






")

")



